
| Hjem > Slåttemyra > Om myr, torv, torvmoser, grunnvann, torvmassivenheter, myrtyper, myrstrukturer, gradienter og indikatorarter |
Om myr, torv, torvmoser, grunnvann, torvmassivenheter, myrtyper, myrstrukturer, gradienter og indikatorarterMyr kan defineres som et område med torv og en hydrofil vegetasjon som selv kan danne torv gjennom ufullstendig nedbrytning. Torv består av organisk materiale produsert på stedet i et område mettet av vann og med underskudd på oksygen. |
Litt om grunnleggende myrterminologi fra 1970- og 1980-talletUtdrag fra en avhandlig av Tor Øystein Olsen fra 1975: A. Definisjon av myrHovedkilder: Begrepet myr defineres på mange forskjellige måter. Resultatene blir forskjellige om utgangspunktet er biologisk, geologisk, geografisk eller kombinasjoner av disse. Biologisk er myra voksested og substrat for en bestemt vegetasjonstype, geologisk er myra en lagrekke av torv, mens den geografisk sett er et landområde. Det er vanligst å definere myr etter en geografisk term. Flatberg (1970) har definert myr på følgende måte: "Med begrepet en myr vil jeg her forstå et område med torv og en hydrofil vegetasjon som selv kan danne torv." Det norske myrselskapet benytter betegnelsen myr på et område "når den mineralske undergrunn er dekket av et torvlag av en viss tykkelse". Når torvtykkelsen brukes i definisjonen, er det mer naturlig å bruke betegnelsen torvmark (geologisk term). Torv kan defineres på følgende måte: "[…] en genom vattenindränkning konserverad jordart, bildad av rester av på platsen växande makroskopisk vegetation." (Sjörs, Hugo 1948. Myrvegetation i Bergslagen. – Acta Phytogeographica Suecica 21: 1–299. side 43) B. Inndeling etter dannelsenHovedkilde: A. Moen 1973: s. 175 Akkumulering av organiske jordarter (torv) skjer når produksjonen av organisk stoff er større enn nedbrytingen. Etter dannelsesmåten deles myrene i gjenvoksningsmyr og forsumpningsmyr. Gjenvoksningsmyrene. Disse har begynt som organiske avsetninger i vatn. Forsumpningsmyrene. Der har torvdannelsen funnet sted på fastmark. Størstedelen av myrarealet i Norge er dannet på denne måten. C. Ombrotrofe og minerotrofe myrområderHovedkilde: K.I. Flatberg 1970: s. 29 Ved klassifisering har jeg søkt å skille mellom ombrotrofe og minerotrofe myrområder. Ombrotrofe myrområder: Minerotrofe myrområder: D. Hydrologisk inndelingHovedkilder: K.I. Flatberg 1970: s. 24–26, A. Moen 1973: s. 175–176. Hydrologiske definerte myrtyper De myrene som i tillegg også får tilførsel av vatn som har vært i kontakt med mineraljorda (minerogent vatn), minerogene myrer (= jordvannsmyrer), deles videre etter den måten myrene får sitt grunnvatn på:
E. Geografiske begreperHovedkilder: K.I. Flatberg 1970: s. 30–34, A. Moen s. 1973: 176. Alle de hydrologiske definerte myrtyper nevnt i kapittel D, kan finnes innenfor ett og samme myrkompleks. Et myrkompleks er et hydrologisk mer eller mindre vel avgrensa landskapsøkologisk system sammensatt av flere voksestedstyper. En voksestedstype kan defineres som alle myrøkosystemer med overensstemmende ernæringsforhold. Ved undersøkelser av myrkompleksene er det da naturlig å studere mindre deler for seg; myrelement. Pr. definisjon er et myrelement det største myrområde som har en noenlunde enhetlig struktur, eller der strukturenes fordeling er noenlunde lik over hele området. I motsetning til voksestedstypene er altså myrelementene reelle naturobjekter. Myrelementene bygger opp myrkompleksene. Generelt kan man si at man har tre økologiske kategorier av myrelementer:
Myrkompleksene har de samme kategoriene. I en grov klassifisering av et myrkompleks, er det den dominerende del av et kompleks som blir avgjørende for komplekset. Innenfor myrelementene kan det være store forskjeller bl.a. mellom tørre og våte partier, og elementene består av strukturer. Myrstruktur: Forekomstflekker av plantesamfunn med en spesiell biotop. Oversikt over noen myrstrukturer: Høljer: Ombrotrofe forsenkninger av variabel grad, og som i det minste i visse perioder av året har en høy grunnvannstand, over eller nær overflata. Som høljeunderstrukturer kan man betrakte termene fastmatter, mjukmatter og løsbunner, sjøl om de egentlig er definert på floristiske kriterier. Flarker: Avgrensa bløte partier, med en glissen, svakt torvdannende, minerotrof vegetasjon, og med avvekslende oppdemmende tørrere partier. Dråg: Smale minerotrofe myrstriper mellom ombrotrofe områder og betinga av soligen markfuktighet. Strenger: Lange, smale forhøyninger som virker demmende på et myrkompleks – vekslende med høljer eller flarker. Andre strukturer er (jfr. egen liste): b) minerotrofe: flarkgjøler, sumper, myrbekker, kilder F. Inndeling etter vegetasjonenFattig–rik, myrflate–myrkant og løsbunn–tue Hovedkilder: K.I. Flatberg 1970: 34–40, A. Moen 1973(b): 180, A. Moen 1973(a): 99–103 Innenfor det enkelte myrkompleks kan det også være store forskjeller i vegetasjonen, og det skilles mellom tre lokale hovedgradienter. Vegetasjonsgradientene henger sammen med variasjonen i økologiske forhold (økologiske gradienter).
Les mer her: NTNU Vitenskapsmuseet: Myr: forskning, forvaltning og overvåkingNTNU Vitenskapsmuseet. Myr: forskning, forvaltning og overvåking Fra Moen, A. & medarbeidere 1983. Myrundersøkelser i Nord-Trøndelag i forbindelse med den norske myrreservatplanen. – K. Norske Vidensk. Selsk. Mus. Rapp. Bot. Ser. 1983-1: 1–160. Fordeling av arter langs de økologiske gradientene – økokliner (tidligere vegetasjonsgradienter)Oversikt over vegetasjonsenhetene for myr, høgstarrsump og kilde
Fordeling av noen typiske myrplanter på tue, fastmatte, mykmatte og løsbunn for nedbørsmyr, fattigmyr og rikmyr
Oversikt over myrartenes fordeling på ombrotrof myr
Fordeling av arter i myrvegetasjon langs fattig–rik-gradienten
Fordeling av arter i myrvegetasjon langs vegetasjonsgradienten tue–løsbunn
Fordeling av arter langs vegetasjonsgradienten myrflate–myrkant
(Side 48.) "Fastmatte av fattigmyr er den dominerende vegetasjonsenhet (jfr. fig. IV, Kap. II, 6 som viser enhetene). Det fins flere utforminger av enheten, men som regel er arter som bjønnskjegg (Scirpus caespitosus = Trichophorum cespitosum cespitosum), torvull (Eriophorum vaginatum), duskull (E. angustifolium), blåtopp (Molinia) og flaskestarr (Carex rostrata) blant de vanligste artene. Rome (Narthecium) dominerer også store myrareal. De nevnte artene har vært viktige for produksjonen av fór på myrene, og store myrareal har vært nyttet som slåtteareal helt fram til siste krig. Intermediær, rik og ekstremrik vegetasjon fins spredt i fylket, og vanligvis utgjorde de rike myrene de beste slåttemyrene." Les om Slåttemyra og slåttemyrer her. Les også: Myra. Ei populærvitskapleg framstilling av myr i NiN-systemet.Utdrag fra rapporten"Myra er den einaste terrestriske naturtypen som bygger sitt eige substrat, torv. Vi nyttar dette særdraget når vi definerer myr som eit landområde med vegetasjon som krev fukt, og som dannar torv. Tørrstoffet i torva er i stor grad restar av døde plantar, hos oss er mosane svært viktige, men og karplanter bidreg til å lage torv. Blant mosane er torvmosane (Sphagnum spp.) torvbyggarane framfor nokon. Eit høgt vassnivå hindrar nedbryting av organisk materiale, og er avgjerande for danning av myr. På eit overordna nivå er det klima, topografi og eigenarten til mineraljorda eller berggrunnen som avgjer kor det blir danna myr, og kva for type myr som veks fram. Myrnatur kan delast inn på fleire måtar, og i NiN er det inndeling etter vegetasjon samt den hydromorfologiske karakteriseringa som er sentral. Utskifting av artar langs økologiske gradientar ligg til grunn for inndeling i hovudtypar og grunntypar, det vil seie at det er inndeling etter vegetasjon som blir brukt i typesystemet. Dei aktuelle hovudtypane (tal grunntypar i parentes) er: V1 Åpen jordvannsmyr (32), V2 Myr- og sumpskogsmark (8), V3 Nedbørsmyr (7), V4 Kaldkilde (9), V8 Strandsumpskogsmark (3), V9 Semi-naturlig myr (3), V11 Torvtak (2), V12 Grøftet torvmark (3) og V13 Ny våtmark (8). Dei viktigaste økologiske gradientane på myr har tradisjonelt vore kalla fattig–rik, mjukmatte–tue, og myrkant–myrflate. I tillegg kjem påverknad frå kjelder inn som ein viktig gradient. I NiN blir nemningane kalkinnhold (KA), tørrleggingsvarighet (TV), myrflatepreg (MF), og kildevannspåvirkning (KI) brukt om desse gradientane, og dei er døme på lokale komplekse miljøvariabler (LKM)." IndikatorarterSe vedlegg om:
Les om Våtmarkssystemer i NiN. Torvmarksmassiv i NiN"Torvmarksmassiv brukes for å beskrive myr som landform, og beskriver torvforekomstens egenskaper, ikke myrøkosystemet på torvoverflata. Typeinndelingen i Torvmarksmassiv er resultatet av de økologiske forholdene som har virket på myrdanning og myrvekst over tid." Kjell Ivar Flatberg: Norges torvmoser
Klikk på bildet, og kjøp boka som E-bok, eller les om de 57 artene med torvmoser hos Artsdatabanken: Torvmoser hos Artsdatabanken: Flatberg KI (2015). Torvmoseslekta Sphagnum L. Les også: https://www.artsdatabanken.no/Pages/205793/Hvor_i_myra_ Utdrag fra boka:Torvmosenes voksested (naturtyper og habitat)(Side 28–45.) […] "Under våtmarkssystemer er det funnet mest hensiktsmessig å skille ut Myr og Kilde og kildemark som egne og to separate natursystemenheter." […] "Våtmarkssystemer: Kilde og kildemark (Min anmerkning: Geogen myr er synonymt med minerogen myr.) Våtmarkssystemer (VåS)[…] "Torvmosene er mer enn noen annen plantegruppe med på å danne, utforme og visualisere vårt myrlandskap, og er de langt viktigste bidragsyterne til torv- og myrdannelse. Videre er torvmosene viktige indikatorarter for vitenskapelig klassifisering av ulike myrtyper både lokalt og regionalt. Det er derfor lagt vekt på å definere, avgrense og karakterisere ulike enheter og grunntyper i norsk myrnatur. Klargjøring av vitenskapelig myrterminologi inngår som en viktig del av dette. Omtale av viktige økologiske og biologiske prosesser som bestemmer og påvirker lokal og regional utbredelse for de ulike torvmosene som vokser der, inngår også." […] Myr"Myrklassifisering, prinsipper og terminologi brukt her bygger på skandinavisk tradisjon. Grunnlaget er den svenske såkalte 'Uppsala-skolen' på 1940–50-tallet, med sentrale myrforskere som H. Sjörs og G.E. Du Rietz. Denne skolen har siden 1970 også vært retningsgivende for klassifisering av norsk myr og myrvegetasjon, både på forskningssiden og for anvendt, forvaltningsrettet faglig utredningsarbeid, blant annet i arbeidet med utarbeidelse av den norske myrreservatplanen. Viktige referansearbeider i denne sammenheng er Sjörs (1948b, 1950, 1952), Du Rietz (1949, 1954), Malmer (1962), Flatberg (1970), Moen (1970, 1973, 1985, 1990, 1995), Fransson (1972), Økland (1989a, b; 1990a, b, c), Rydin et al. (1999). Grunnleggende for inndeling og klassifisering av myr og myrvegetasjon i denne skolen har vært studier og beskrivelse av flora og vegetasjon i forhold til variasjon langs lokale økologiske gradienter (økokliner). En rekke av opplysningene nedenfor bygger på denne litteraturen, men mye bygger også på egen felterfaring og upublisert kunnskap, til dels på usitert spesiallitteratur. Myr omfatter landområder med torv av varierende tykkelse og en vegetasjon av fuktighetskrevende og/eller fuktighetstolererende planter som gjennom ufullstendig nedbrytning på stedet danner torv. Torv er en jordart som består av ufullstendig nedbrutte plantedeler, og der vekten av organisk materiale utgjør mer enn 30 %. Torvmark omfatter våtmark med minst 30 cm torvdybde, og er derfor arealmessig et snevrere begrep enn myr. En forutsetning for torv- og myrdannelse er et grunnvannsspeil som ligger i eller nær markoverflaten gjennom mesteparten av året og forårsaker god markfuktighet og oksygenfattige forhold i, og nær markoverflaten. MyrterminologiMyrbegrepet har i fennoskandisk myrforskning oftest vært knyttet opp mot myr og skogsmyr, mens sumpskog har vært holdt utenfor og blitt oppfattet som en fuktekstrem utforming av fastmarksskog. Definisjonene og omtalene nedenfor avspeiler denne tradisjonen, men myrbegrepet omfatter også myrskog i vid betydning, dvs. at sumpskoger er inkludert. Med grunnlag i grunnvannets opprinnelse og kjemiske sammensetning deles myr i to hovedtyper:
De norske betegnelsene på disse to hovedmyrtypene er henholdsvis nedbørmyr og jordvannsmyr. De tilsvarende økosystembegrepene som inkluderer de biologiske systemene og artene som inngår, er henholdsvis ombrotrof og minerotrof myr. Det innebærer for eksempel at et bestemt myrområde har ombrotrof eller minerotrof vegetasjon. Navnene nedbørmyr og jordvannsmyr brukes her ofte i begge betydningene. De geogene myrene kan deles inn i:
Helningsmyrer dominert av fastmattevegetasjon, og med mer enn 3° helning, blir ofte klassifisert som bakkemyrer. Disse er mest vanlig i MeBo og NoBo. Ingen torvmoser er i utbredelse eksklusivt bundet til limnogen myr og bakkemyr. I klassifiseringen og omtalen av de enkelte torvmoseartenes voksested er det ikke skilt mellom flatmyr, flommyr og helningsmyr. Alle norske fastlandsarter av torvmoser kan forekomme på myr. De fleste artene har også sine hovedforekomster der, men bare et begrenset antall arter kan sies å være tilnærmet eksklusive myrarter. Slike arter er f.eks. glasstorvmose Sphagnum angermanicum, lapptorvmose S. subfuvlum subsp. subfulvum, flarktorvmose S. jensenii, trøndertorvmose S. troendelagicum og huldretorvmose S. wulfianum. De fleste artene har også forekomster i andre våtmarks- og fastmarkssystemer. I tillegg inngår mange torvmoser på myr som er like vanlig eller vanligere i fastmarkssystemer (fastmarksskog, hei). Slike arter er f.eks. grantorvmose S. girgensohnii, tvaretorvmose S. russowii, furutorvmose S. capillifolium, lyngtorvmose S. quinquefarium, heitorvmose S. strictum. Jordvannsgrensen, jordvannsindikatorgrensen Blant karplantene er de fleste starrartene som forekommer på myr, gode jordvannsindikatorer. Gode eksempler er flaskestarr Carex rostrata og trådstarr C. lasiocarpa. Det er bare sveltstarr C. pauciflora (i Vest-Norge), dystarr C. limosa og snipestarr C. rariflora som kan forekomme på ombrogen myr. Det eksisterer knapt arter av moser, lav og karplanter som er eksklusivt bundet til ombrogen myr, men mange har sitt viktigste voksested der. De enkelte artenes forekomst på ombrogen og geogen myr varierer regionalt, og jordvannsindikatorgrensen er derfor et relativt begrep. Noen torvmoser oppfører seg forskjellig i vest og øst i deres utbredelsesområde. Dette er korrelert til seksjonsgradienten fra StOs til SvKo, og skyldes primært sterkere tilførsel av havsalter gjennom nedbør, dogg og vind i havnære områder. En kan da få inn arter på ombrogen myr i de sterkest oseaniske seksjonene som er jordvannsindikatorer i mindre oseaniske og mer kontinentale seksjoner. Vortetorvmose Sphagnum papillosum er en vanlig ombrogen myrart i kystområdene i Vest- og Midt-Norge og Nordland, men er på indre deler av Østlandet en god jordvannsindikator. Dette gjelder også grantorvmose S. girgensohnii og tvaretorvmose S. russowii som i Vest- og Midt-Norge er vanlig i ombrogen åpen myrkant, men som er jordvannsindikatorer mange steder på Østlandet. Bjørnetorvmose S. lindbergii er en vanlig ombrogen art i boreale områder, men er på sørlige forekomster i BoNe på Østlandet oftest jordvannsindikator. I StOs langs vestkysten vokser horntorvmose S. auriculatum i høljegjøler, blanktorvmose S. subnitens subsp. subnitens og heitorvmose S. strictum i ombrogen tuevegetasjon, og fløyelstorvmose S. molle i ombrogen fastmattevegetasjon, mens de ellers er jordvannsindikatorer. Broddtorvmose S. fallax er gjennom mesteparten av landet en god jordvannsindikator som mangler på ombrogen myr. Men enkelte steder på Sør- og Sørvestlandet er den også funnet med spredde forekomster i ombrogene mykmatter. Dette er trolig et resultat av langtransporterte luftforurensninger og de høye våtavsetningene av nitrogen som en har hatt over lengre tid i denne delen av landet. (Min anmerkning: Fastmarksvanngrensen er synonymt med jordvannsgrensen.) Økokliner (økologiske gradienter)Lokalt varierer flora og vegetasjon på myr først og fremst i forhold til to basis-økokliner og en kompleksøkoklin:
[…] "I denne boken brukes betegnelsen gradient (lokale og regionale gradienter), som er anvendt i både økologisk og floristisk/vegetasjonsmessig betydning. De lokale gradientene er:
Denne navnebruken er i samsvar med skandinavisk tradisjon innenfor myrforskning, men tar her også høyde for myrskog som en del av spennet i kompleksgradienten 'myrflate–myrkant'. 'Tørr–fuktig'-gradienten
På ombrogen myr skiller en tradisjonelt mellom tuer og høljer, som utgjør myrstrukturer. Høljene er videre delt inn i fastmatter, mykmatter og løsbunn, mens en mangler en tilsvarende fellesterm på geogen myr. I denne boken brukes hølje i betydningen all torvmosedominert myr som ligger under tuenivå på ombrogen og fattig geogen myr. Gjøler er åpne, sekundære vannansamlinger som er dannet etter at myra ble til. Høljegjøler er slike vannansamlinger på ombrogen myr, og flarkgjøler på geogen myr. Flarker er mykmatte- og løsbunnområder på geogen eller svakt soligen myr som oftest er omgitt og oppdemt av langstrakte fastmatte- eller tuestrenger. Myrtjern er åpne, primære vannansamlinger som var til stede før myra ble dannet, og som er omgitt av oftest geogen myr, og oftest er under gradvis gjenvoksning fra kantene (gjenvoksningsmyr). Variasjonen innenfor de ulike gradientene avspeiles av artssammensetningen, og torvmoser er sentrale indikatorarter. Grensen mellom tuemyr og mattemyr kan på mange myrer defineres gjennom nedre røsslynggrense. Røsslyng Calluna vulgaris og flere andre lyngarter har rotmykorrhiza (samliv med sopp) og tåler ikke vanndekning av røttene. De ulike mattenivåene er ikke alltid klart botanisk adskilte. Flytemattemyr er her utskilt som et eget trinn. Det kan betraktes som en ekstrem vannmetningsutforming av mykmatte med torvmoseskudd som vokser vanndekte gjennom hele eller store deler av sommerhalvåret, men som i artssammensetning ikke er klart adskilt fra mykmattemyr. Løsbunnmyr er ikke tatt med som eget trinn da en her i liten grad finner torvmoser, og ingen torvmosearter viser preferanse for slik myr. 'Fattig–rik'-gradienten
Variasjonen innenfor gradienten kan karakteriseres gjennom artsammensetningen, og torvmoser er viktige indikatorarter. Kalkmyr (ekstremkalkrik myr) mangler i det store og hele torvmoser, og er i denne boken ikke skilt ut som eget forekomsthabitat for torvmoser. Men enkelte arter kan nå og da ha spredde forekomster også på ekstremkalkrik myr, da gjerne som innblanding blant 'brunmoser' som myrstjernemose Campylium stellatum og brunmakkmose Scorpidium cossonii. Vritorvmose Sphagnum contortum er det beste eksemplet på en slik torvmoseart, men også rosetorvmose S. warnstorfii, lapptorvmose S. subfulvum subsp. subfulvum og beitetorvmose S. teres inngår nå og da i kalkmyr. Innenfor kalkfattig myr kan det skilles ut to typer: ekstremkalkfattig og moderat kalkfattig myr, men skillet er ofte glidende og vanskelig å definere gjennom artssammensetningen. Gjennom måling av surhetsgraden (pH) i myrvannet kan en få et indirekte mål på graden av kalkinnhold i de ulike typene.
Næringstilførselen til ombrogen myr varierer. På åpen ombrogen myr vil tuene langt på vei få all sin næringstilførsel fra nedbøren som faller på stedet. De underliggende forsenkningene (høljene) vil oftest ha noe høyere næringsinhold og pH. Dette avspeiler bedre næringstilgang, dels gjennom 'avrenning' fra tuene, dels gjennom den vanligere forekomsten av nitrogenfikserende blågrønnalger i høljene, både som frittlevende og som 'beboere' i torvmosenes hyalinceller. Tuetorvmosenes større evne til kationutveksling (se Torvmosenes egenskaper), gir også lavere pH enn i høljene. Tuene i ombrogen myrskog vil også ofte ha et noe høyere næringsinnhold og pH enn på myrflate. Dette skyldes først og fremst ekstra tilført næring utenom nedbøren i form av strøfall fra trærne som vokser der, men også andre kanteffekter. 'Myrflate–myrkant'-gradienten Eksposisjonsforholdene er viktige for forekomst og utseende hos torvmoser. Skygge skapt av trær og busker – delvis også av lyng og andre dvergbusker – påvirker i høy grad skudd-og hodefarge, vekstform, lengdevekst/produksjon og blad- og cellemorfologi hos mange torvmoser. Denne eksposisjonsgradienten blir her omtalt som 'sol–skygge'-gradienten. NisjedifferensieringTorvmoser viser en utpreget grad av nisjedifferensiering i forhold til lokale økologiske gradienter, dvs. at de som regel bare er i stand til å vokse innenfor en avgrenset del av det totale økoklinspennet, når en ser bort fra konkurranse med andre arter. Dette er den fundamentale nisjen, som avspeiler artenes fysiologiske toleranse. Den realiserte nisjen er den faktiske nisjen når arten i konkurranse med en eller flere andre arter blir ekskludert fra deler av sin fundamentale nisje. Dette er på myr oftest andre torvmosearter. Den realiserte nisjen til en bestemt torvmoseart er derfor oftest smalere enn den fundamentale." […] Myrnaturtypene"Ombrogen myrskog Én grunntype er skilt ut i forhold til 'tørr–fuktig'-gradienten:
Ombrogen åpen myrkant To grunntyper er skilt ut i forhold til 'tørr–fuktig'-gradienten:
Ombrogen myrflate Ulike torvmosearter dominerer bunnsjiktet på ombrogen myrflate, særlig i fastmatter og mykmatter/flytematter, men mange steder inngår eller dominerer også torvmoser på tuemyr. Særlig rusttorvmose Sphagnum fuscum er på tuene en karakteristisk art. Men ingen torvmosearter er funnet bare på ombrogen myrflate. Torvmosene er oftest klart nisjefordelt i forhold til 'tørr–fuktig'-gradienten. Fire grunntyper er skilt ut i forhold til 'tørr–fuktig'-gradienten:
Geogen myrskog
Geogen åpen myrkant I forhold til 'tørr–fuktig'-gradienten forekommer alle myrtyper fra tue- til mykmattemyr, med mest fastmattemyr. Langs 'fattig–rik'-gradienten finnes myrtyper fra kalkfattig til ekstremkalkrik. Geogen åpen myrkant er rikere på torvmosearter enn noen andre myrtyper. Mange av jordvannsindikatorene blant torvmosene har sine hovedforekomster her. Noen få torvmosearter har også sin utbredelse begrenset til slik myr, f.eks. trøndertorvmose S. troendelagicum. Ni grunntyper er skilt ut i forhold til 'fattig–rik'- og 'tørr–fuktig'-gradientene:
Geogen myrflate Tolv grunntyper er skilt ut i forhold til 'fattig–rik'-gradienten og 'tørr–fuktig'-gradienten:
[…] (Side 82–93.) Torvmosenes egenskaper"Torvmosenes suksess som myr- og torvdannere er betinget av flere forhold. De overordnede faktorene/forholdene er:
Disse forholdene er betinget og regulert av en rekke egenskaper og prosesser hos torvmosene: De kan ta opp og holde på store mengder vann. Dette skyldes først og fremst bladenes spesielle celleoppbygning der de store og tomme vannlagringscellene (hyalincellene) utgjør det meste av bladvolumet, men også gjennom greinenes og bladenes morfologi og arrangement som muliggjør effektiv kapillær transport og holding av vann, og som ytterligere fremmes av skuddenes matte- og puteaktige vekstform. Artene har ulik evne til å motstå uttørking (uttørkingsresistens), holde på vann (vannholdningskapasitet) og til å tåle tørke (tørketoleranse), dvs. evnen til å unngå og tåle uttørking varierer artene imellom. Effektiv kapillær transport av vann i skuddene i kombinasjon med høy vannholdningskapasitet gir høy uttørkingsresistens. Tueartene har større vannholdningskapasitet, tørketoleranse og uttørkingsresistens enn høljeartene, og gjenopptar effektiv fotosyntese og livsfunksjoner raskere etter tørke. Den tidligere påstanden om at høljearter har større tørketoleranse enn tuearter, dvs. at de tåler å stå uttørket i lengre tid enn tueartene, er trolig ikke riktig (Hajek & Beckett 2008). De fleste torvmosene tåler å stå vanndekte over lengre perioder. Torvmosene har uvanlig høy kationbyttekapasitet. Dette muliggjør effektiv utnyttelse av lave konsentrasjoner av kationer fra vannoppløsninger, f.eks. av Ca++-ioner. Kationbyttestedene er COO--grupper (karbonylgrupper) i uronsyrer (galakturonsyre og 5-keto-D-mannuronsyre, 5KMA) som er lokalisert i bladenes cellevegger. Nye kationbyttesteder (trolig som COOH) dannes fortøpende når mosen vokser. Med en fellesbetegnelse kalles de pektinlignende polysakkaridene i celleveggene hos torvmoser der uronsyrene er lokalisert, for sphagnan. Den høye kationbyttekapasiteten gjør at mange torvmoser kan ernære seg fullt og helt gjennom opptak av nødvendige mineralnæringsstoffer fra regnvann og atmosfæren, som på nedbørmyr. Uronsyrene utgjør ca. 17 % av tørrvekten hos mykmattearten vasstorvmose Sphagnum cuspidatum og ca. 21 % hos tuearten rusttorvmose S. fuscum. Dette gjør at tueartene på myr har høyere kationbyttekapasitet enn matteartene. Også innenfor en og samme art vil kapasiteten øke med minkende fuktighet på voksestedet. Ved aktiv vekst byttes vannoppløste kationer i cellenes næromgivelser (f.eks. Ca++-ioner og andre kationer) ut ved at H3O+-ioner i uronsyremolekylene frigjøres til vannet omkring, bivalente (++) ioner før monovalente (+). Torvmosene bidrar derfor i sterk grad til å forsure sitt eget miljø, og er den viktigste årsaken til myrenes gjennomgående sure vann- og torvmiljø. Dette avspeiles for eksempel i at pH i grunnvannet på nedbørmyr oftest ligger omkring 3,7–4,3, mens regnvann vanligvis har pH på ca. 4,8–5,1, dvs. regnvannet er gjort surere gjennom kationbytte. De fleste torvmosene vantrives i vannmiljø med for høy pH der for mange kationer bindes opp, og i omgivelser med for lav pH der alle andre kationer byttes ut fra ionbyttestedene. Kombinasjonen av høye konsentrasjoner av Ca++ og høy pH i grunnvannet er skadelig til dødelig for de fleste myrtorvmoser. Dette skyldes trolig at Ca++-ionene vil bli preferert i ionbyttet og 'fyller' opp ionbyttestedene på bekostning av andre kationelementer som er nødvendige for mosens eksistens og vekst. pH inne i cellene påvirkes i liten grad av ionbytteprosessen, og pH er derfor normalt høyere enn i vannet utenfor. Myrtorvmoser har uvanlig lav nedbrytningshastighet. Dette skyldes lavt innhold av tilgjengelig nitrogen i skuddene (< 1 %), de sure omgivelsene skapt av torvmosene selv, det vannmettede og oksygenfattige nærmiljøet, og at celleveggenes sphagnan brytes ned sakte og går over i vannløselig form først lang tid etter at mosen er død. Det er vist at sphagnan binder opp og deaktiverer enzymer som påvirker mikrobiell nedbrytning av organisk materiale. Sphagnan binder også effektivt opp ammonium NH4+ fra næromgivelsene, og bidrar dermed til at næringsgrunnlaget for nedbrytningsorganismer (bakterier, sopp) blir redusert. Det har lenge vært registrert at torvmoser har antimikrobielle og konserverende egenskaper. Egenskapene skulle være knyttet til 5-KMA-forbindelsen i sphagnan som var i stand til å deaktivere viktige enzymer som mange mikroorganismer er avhengige av. Den tidligere antagelsen om at torvmoser inneholder antimikrobielle fenolforbindelser (ofte kalt for sphagnol), har vist seg å være uriktig. Slike fenolforbindelser (særlig 'Sphagnum-syre) kan riktig nok bidra til forsuring og lavere nedbrytning av plante- og dyrerester, men konsentrasjonene de opptrer i, er for lave til å forklare torvmosenes antibakterielle egenskaper (Mellegård et al. 2009). Annen nyere forskning viser også at en må sette et spørsmålstegn ved de antimikrobielle egenskapene knyttet til sphagnan, bortsett fra å motvirke bakterievekst gjennom senkning av pH i omgivelsene (Stalheim et al. 2009). På den andre side har undersøkelser vist at torvmoser har samfunn av ekto- og endohydriske bakterier, særlig innenfor slekten Burkholderia, som produserer soppdrepende (antifungale) forbindelser (Opelt et al. 2007a). Disse kan være med på å forklare – eller delforklare – torvmosenes antimikrobielle egenskaper. Over årtusener frigjøres etter hvert sphagnan fra de døde torvmoserestene i torva til vannet omkring. Her danner det sammen med ammonium og aminosyrer en vannholdig humus, som virker garvende og konserverende på proteiner (Painter 1995). Hud på dyr- og menneskekropper kan på denne måten omdannes til læraktige strukturer gjennom en garveprosess, som går under navnet Maillards reaksjon (Painter 1998). Godt oppbevarte, flere tusen år gamle lik i myrer, som er funnet flere steder i Nordvest-Europa, vitner om dette. En kan i de fleste norske myrer også finne forbausende intakte røtter, stubber og trestammer i to adskilte lag nedover i torva. De er rester etter trær som vokste der i boreal og suboreal tid, henholdsvis ca. 7400–6200 og 3000–500 f.Kr. Først og fremst gjelder dette furu Pinus sylvestris, som forsvant fra myra i fuktigere og kaldere perioder i etteristiden. Pollenkorn, sporer og frø opprinnelig avsatt på myroverflata, blir også konservert og oppbevart på samme måte. Vegetasjonshistoriske studier med pollenanalyse fra torvprøver i ulike lag nedover i myr baserer seg på dette. Konserveringen av pollenkornene gir så lite omdanning at en fremdeles kan bestemme hvilken art eller artsgruppe de tilhører. Torvmosene har en effektiv vegetativ vekst og formering som gir tette matter og puter som kan dekke store områder. Torvmoser beites ikke av dyr. Torvmosenes vekst og produksjonLengdevekst og produksjon hos torvmoser varierer både innenfor og mellom arter i et bestemt geografisk område med samme makroklima, for eksempel på en og samme myr. Myrtorvmoser viser avtagende lengdevekst med minkende fuktighet langs 'tørr–fuktig'-gradienten. Det innebærer for eksempel at rødtorvmose Sphagnum rubellum kan ha en årlig tilvekst som varierer mellom ca.10 mm når den vokser i tuer, til ca. 70 mm i mykmatter, ifølge en undersøkelse gjort på De britiske øyer (Clymo 1970). På en myr på Sørlandet er vortetorvmose S. papillosum registrert med en årlig lengdetilvekst på ca. 30 mm i mykmatte og noe i underkant av 10 mm i fastmatte (Pedersen 1975). På samme myr ble broddtorvmose S. fallax registrert med en årlig lengdevekst på ca. 120 mm i mykmatte. Generelt har fast- og mykmattearter større lengdetilvekst enn tuearter. Tuearten rusttorvmose S. fuscum er på en myr i Finland målt med en årslengdevekst på 8–18 mm, mattearten vortetorvmose S. papillosum 25–32 mm, og mykmattearten skartorvmose S. riparium til 35–54 mm (Lindholm & Vasander 1990). I flytematter på myr kan en noen ganger finne torvmoseskudd som er meterlange, og som er resultat av mange års vekst. Tilsvarende kan en på overhengsmatter på bergvegger finne puter av torvmoser med levende skudd på 20–30 cm. Netto primærproduksjon (NPP) blir ofte brukt som mål på produksjonen, og kan uttrykkes gjennom tørr biomasse som blir dannet pr. kvadratmeter pr. år. Primærproduksjonen i en torvmosematte/-pute synes generelt å ligge innenfor en spennvidde på 100–600 g/m2/år (Clymo & Hayward 1982), og er normalt lavest på tuer og høyest i fast- og mykmatter (Gunnarsson 2005). Men den behøver ikke å være korrelert med lengdeveksten når en sammenligner ulike arter. På en undersøkt myrlokalitet på kysten i British Columbia, Kanada, hadde rusttorvmose S. fuscum på tue en årsvekst på 12,5 mm og en årsprodukasjon på 365,3 g/m2, mens mykmattearten bjørnetorvmose S. lindbergii hadde en årsvekst på hele 59,6 mm, men lavere årsprodukjson på 359,9 g/m2 (Asada et al. 2002). Økt skygge har påvist økt lengdevekst, men synes i mindre grad å påvirke produksjonen (Clymo & Hayward 1982). Dette forholdet kompliseres trolig også av samvirkende effekter av 'sol–skygge'- og 'tørr–fuktig'-gradientene. Det er mer uklart hvordan produksjonen påvirkes i forhold til 'fattig–rik'-gradienten. Regionalt og globalt er årsproduksjonen av torvmoser på myr først og fremst regulert av 'fotosyntetisk aktiv stråling' (PAR) gjennom vekstsesongen (Loisel et al. 2012), og ulike variabler knyttet opp mot temperatur og sommervarme, for eksempel uttrykt gjennom gjennomsnittlig årstemperatur (Gunnarrsson 2005), vekstsesonglengde og sommertemperatur. I praksis innebærer det at produksjonen på den nordlige halvkule minker med økende breddegrad og høyde over havet, dvs. når en går fra nemoral bioklimatisk sone, via boreale til arktiske soner, og fra lavlandet til alpine soner. En har også indikasjoner på at myrtorvmoser har lavere produksjon i områder med kystklima enn med innlandsklima. Dette er korrelert med høyere PAR og vekstsesongtemperaturer med økende kontinentalitet og minkende oseanitet. Jevnt over tettere skydekke i kystområder kan være en viktig årsak, men et mer stabilt snødekke i innlandsområder kan også gi bedre beskyttelse mot frostskader på torvmosehodene enn det manglende eller ustabilt snødekke i kystområder gir, f.eks. i StOs langs Norges vestkyst. I tillegg til klimatiske faktorer vil også ulike næringsstoffer påvirke vekst og produksjon, for eksempel ulik belastning av nitrogen (N) gjennom menneskeskapt virksomhet og luftforurensninger (Dorrepaal et al. 2003). Forholdet mellom fosfor (P) og nitrogen (N) som begrensende vekstfaktor på ombrogen myr blir også påvirket av dette. I nordboreale områder i nordlige Skandinavia er veksten og produksjonen ofte nitrogenbegrenset, mens den i nemorale/boreonemorale områder i sør med høye, menneskeskapte N-avsetninger er fosforbegrenset (Aerts et al. 1992). Gunnarsson og Rydin (2000) har vist at nitrogengjødsling reduserer torvmoseproduksjonen på svenske nedbørmyrer. Tilsvarende vil også nedbørens innhold av 'sure' komponenter ('sur nedbør') og forurensende og toksiske elementer, f.eks. svovelforbindelser, påvirke produksjonen negativt. Torvmosene som myrutformerTorvmoseartenes habitatpreferanser og ulike økofysiologiske og biokjemiske egenkaper, skaper langt på vei det kuperte overflaterelieffet av tuer og høljer en finner på de fleste åpne, treløse nedbørmyrer. Tuevoksende torvmoser har mindre vekst og produserer mindre biomasse enn høljeartene. Det akkumuleres likevel totalt sett mer biomasse av døde skuddrester (førne) i tuene enn høljene. Dette skyldes at tueartene produserer førne av annen kvalitet enn høljeartene og er mer motstandsdyktige mot nedbrytning. Tuetorva i det øverste sjiktet inneholder derfor mindre omdannet torvmoseførne enn i høljene, og torvtilveksten blir større. En vet ikke med sikkerhet hvilke forhold som forårsaker dette. Men forklaringen kan ligge i tueartenes høyere innhold av sphagnan og flere forekomster av kation-byttesteder. Tilsvarende forhold gjør seg også gjeldende på torvmosedominert jordvannsmyr med veksling mellom tuer og høljer. Torvmosene er derfor i stand til å skape sitt eget voksested og utforme mikrotopografien med vekslende tuer og høljer. På ombrogen myrflate skjer dette nesten utelukkende gjennom næringstilførsel via nedbøren som faller på stedet. I tillegg er det vist at forekomst av karplanter (særlig lyngarter) er viktig i både dannelse og opprettholdelse av tuer på ombrogen myr (Pouliot et al. 2011a). Som vist av Karofeld og Pajula (2005) påvirker også døde flekker av torvmoser forårsaket av sopp- og algeangrep fordelingsmønstret av tuer og høljer. Myrtuene på nedbørmyr og fattig jordvannsmyr er først og fremst dannet av arter i underslekt Acutifolia, der rusttorvmose Sphagnum fuscum er den vanligste arten. Kysttorvmose S. austinii, kjøtt-torvmose S. medium og vortetorvmose S. papillosum i underslekt Sphagnum kan også være vanlige tuearter. Høljene er først og fremst dannet av arter i underslekt Cuspidata, og de vanligste artene er bjørnetorvmose S. lindbergii, dvergtorvmose S. tenellum, svelttorvmose S. balticum og vasstorvmose S. cuspidatum. Klubbetorvmose S. angustifolium er den eneste Cuspidata-arten som er vanlig på tuer. På intermediær og moderat kalkrik myr er Subsecunda-arter vanlige mattearter. Torvmosene som karbonlagrerTorvmarker dekker globalt ca. 4 x 106 km2 eller ca. 2–3 % av jordas landareal, med hovedforekomstene på den nordlige halvkule. Siden siste istid har de lagret over 600 Gton (= 600 milliarder tonn) karbon (Clymo 1998). Omtrent 30 % av landjordas karbon er lagret i myr og torvmarker (Gorham 1991). Ett gram torv inneholder ca. 0,5 g karbon (Joosten & Clarke 2002). Det intakte myrarelaet ligger i Norge i dag på på ca. 23 000 km2 (Moen et al. 2011), nærmere 3 % av landarealet. Det totale karbonlageret i udyrket myr inklusive torv og vegetasjon i norske myrer ligger på ca. en milliard tonn karbon (Grønlund et al. 2010). I gjennomsnitt dannes ca. 0,5–1 mm med ny torv årlig på våre intakte myrer. Dette innebærer at det tar mellom 1000 og 2000 år å produsere en meter med torv. Totalt årlig tilvekst av torv på norske myrer er på ca. 2,5–5,0 millioner m3. Dette tilsvarer netto CO2-lagring på ca. 1,5–2,5 millioner tonn. I praksis innebærer det at store mengder karbon bindes opp årlig i våre intakte myrer. Myr er også den norske naturtypen som binder mest karbon pr. arealenhet, ca. tre ganger så mye som i skog og jordbruksareal (Grønlund et al. 2010). Det er også den naturtypen som er antatt å være mest sårbar overfor menneskelige inngrep. Torvmoser er en svært viktig bestanddel både i det levende plantedekket og den underliggende torva, og utgjør over halvparten av verdens torvlagre. De har derfor på intakte myrer en svært viktig funksjon som karbonlagrere. Blant plantene står derfor torvmosene helt sentralt for opptak og lagring av karbon, også sett i et globalt perspektiv. Torvmoseslekten lagrer trolig mer karbon enn noen annen planteslekt i verden. Det er vist at økt tilførsel av nitrogen (N) fører til økt vevsnitrogen hos torvmoser, mindre lengdevekst og produksjon, og raskere nedbrytning og økt CO2-utslipp. I områder med høye nitrogenavsetninger i Sørvest-Sverige er det på en undersøkt nedbørmyr funnet at karbonakkumulering nesten ikke forekommer (Gunnarsson et al. 2008). Økt sommertemperatur og høyere årsnedbør påskynder disse negative prosessene i forhold til karbonbalansen. Det vil i praksis kunne innebære at et varmere klima under tilsvarende atmosfæriske nitrogenavsetninger som en har i dag, vil ha en betydelig effekt på evnen hos torvmoser og dermed myr til å lagre karbon (Kivimäki 2011, Limpens et al. 2011). Ved høyere doser av nitrogen som overskrider 'naturens tålegrense' (på nedbørmyr ca. 10 kg N pr. hektar/mål; se Bakken & Flatberg 1995), makter ikke torvmosene lenger å lagre og nyttiggjøre seg tilført nitrogen, som dermed filtreres ned til karplantenes rotsjikt, og veksten avtar (Woodin & Lee 1987, Aerts et al. 1992). Denne tålegrensen har i perioder på slutten av 1900-tallet også vært overskredet i Norge, særlig på Sørvestlandet. Det er vist at slik nitrogenlekkasje på nedbørmyr har ført til endrede konkurranseforhold mellom arter og endret artsammensetning mange steder i Nordvest-Europa. Ofte ser en redusert dekning av torvmoser i bunnsjiktet på bekostning av økt dekning av karplanter i feltsjiktet, f.eks. av blåtopp Molinia caerulea (Hogg et al. 1995). Det er også vist at økt innhold av CO2 i atmosfæren sammen med økte nitrogenavsetninger fører til endringer i artssammensetning og produksjon på torvmosedominerte myrer (Berendse et al. 2001). Torvmoser og metanreguleringSammen med karbondioksid (CO2) og lystgass (N2O) er metan (CH4) den viktigste drivhusgassen. Torvmoser spiller indirekte en viktig rolle i reguleringen av metanutslipp fra myr. Metan dannes på myrer i det underliggende, oksygenfrie torvlaget, som et sluttsteg i en serie av prosesser som bryter ned organisk materiale. Det er anaerobe, såkalte metanogene Archaea (en-cellede, bakterielignende mikroorganismer) som produserer metanen (Parmentier et al. 2011). Metangassen beveger seg oppover i torva og myra og blir der oksidert av aerobe, såkalte metanotrofe bakterier i de øverste og mer oksygenrike lagene av myra. Metanotrofe bakterier er funnet å være til stede både utenpå og inne i torvmosenes hyalinceller (Kolb & Horn 2012). Disse bakteriene er godt tilpasset forholdene i kalde, sure og oligotrofe myrer, med optimale livsbetingelser ved pH 5,0–5,5, moderate temperaturer fra 15–25 oC, og lave konsentrasjoner av mineralsalter i næromgivelsene (Dedysh 2002). Metanotrofe bakterier er også vanlig på myr og våtmarker i permafrostområder på den nordlige halvkule, og kan der oksidere metan ved temperaturer under frysepunktet (Trotsenko & Khmelina 2005). Tjuetre torvmosearter underøkt på en finsk myr hadde alle metanotrofe bakerier (Larmola et al. 2012). Torvmoseskuddene er gjennom vertskap og sin egen fotosyntese en viktig bidragsyter til det oksygenet som de metanotrofe bakteriene bruker for å oksidere metan til CO2. Kabondioksiden går i hovedsak tilbake til torvmoseskuddene for bruk i fotosyntesen og videre vekst. Metanavledet karbon hos torvmoser kan være så høy som 35 % og er med på å forklare det høye karboninnholdet i myrtorv (Parmentier et al. 2011). Overskuddsmetan som ikke blir konsumert av de metanotrofe bakteriene, blir frigjort til atmosfæren. Størst metanoksidering har en imosesjiktet omkring grunnvannsspeilet. I praksis innebærer dette høyere aktivitet i mykmatter av torvmoser enn i fastmatter og tuer (Kip et al. 2012, Larmola et al. 2012). Torvmoser er på denne måten – gjennom å være vertskap for mange metanotrofe bakterier – en viktig kontrollør av metanutslipp til atmosfæren. Metanutslipp fra intakt myr er under nordiske forhold anslått til 8–10 kg (ca. 200 kg CO2-ekvivalenter) pr. dekar/år. Myrdrenering og tilførsel av oksygen til de øverste torvlagene reduserer metandannelse og -utslipp i betydelig grad. Men dette kompenserer på langt nær for det høye karbonutslippet fra omdannet torv på grøftet myr. Bruk av torvmoserBruk av torvmoser i ulike sammenhenger har lang tradisjon, ikke bare i Norge men også i andre land med torvmosevegetasjon. I de siste 10-årene har også nye anvendelsesmåter dukket opp som har skapt ny virksomhet. Isolasjon Konservering av grønnsaker Konservering av fisk Antiseptiske og medisinske produkter Rensing Høsting av torvmoser 'Sphagnum farming' Biologisk overvåking (biomonitoring) Høsting og bruk av torvBruk av brenntorv til oppvarming har lang tradisjon i Norge. Det er kjent tilbake til sagatiden og er omtalt i Snorres kongesaga (EnergiLink 2008). Bruken har vært størst i de kystnære områdene av landet med mangel på vedbrensel. Mer systematisk produksjon av brenntorv begynte i større skala først på slutten av 1500-tallet. På begynnelsen av 1900-tallet ble maskinell innhøsting av torv fra større myrområder vanlig. Dette ga brenntorv både til private husholdninger og industrielle formål. Med tilgang på kull, vannbasert elektrisitet og oljefyring til oppvarming, tok denne virksomheten gradvis slutt i første halvdel av 1900-tallet. Men enkelte steder har tradisjonen med egenprodusert torv til vedfyring holdt seg fram til 1970-tallet. Så sent som under 2. verdenskrig ble det årlig brukt mellom 1,5 og 2 terrawatt-timer (TWh) brenntorv i Norge. Finland er i dag verdens største produsent av brenseltorv, og torvdrevne kraftverk dekker ca. 6–7 % av landets totale energiforbruk (Kirkinen et al. 2007). Torvtaking, torvstikking Strøtorv. Omfatter i dag maskinelt høstet og oppsmuldret myrtorv som i stor skala anvendes til jordforbedring og som dyrkningsmedium i gartner- og hagevirksomhet, med eller uten tilsatt næring. Torvbriketter og torvpotter av ulike slag er mer tilpassede produkter. Utskårne torvblokker brukes også til ulike formål, blant annet i hageanlegg. Bruk av strøtorv i fjøs og stall var tidligere svært vanlig over mesteparten av landet. I blanding med møkk og urin fra dyrene ga det verdifull gjødsel og jordforbedring på åker og eng. Torvuttaket foregikk i det store og hele omtrent som ved torvstikking til brensel, men torvbrikkene var oftest tykkere og ble gjerne lagret til tørk i 'torvhesjer'. Torvbrikkene ble før bruk smuldret opp maskinelt. Ødeleggelse av myrTotalt har Norge i dag ca. 23 000 km2 med intakt myr, mot trolig over 30 000 km2 for 150–200 år siden (Moen et al. 2011). Dette innebærer at ca. 7 000 km2 myr er ødelagt siden den gang, dvs. ca. 23 % av det opprinnelige myrarealet. Myrødeleggelsene skyldes:
Den største bidragsyteren har vært grøfting for skogproduksjon, som har ødelagt over dobbelt så stort myrareal som nydyrkning. Arealmessig har brenn- og strøtorvproduksjon ført til en forholdsvis begrenset myrødeleggelse, i perioden 1919–1960 beregnet til ca. 300 km2 (Moen et al. 2011). Grøfting og oppdyrking av myr til grasproduksjon startet i stort omfang på midten av 1700-tallet, og det ble etter hvert gitt betydelig offentlig støtte til denne virksomheten. Eventyrsamleren P. Chr. Asbjørnsen utga to skrifter om myrutnyttelse som fikk stor betydning (Asbjørnsen 1856, 1868). Denne virksomheten har siden pågått med varierende omfang, og det er fremdeles ikke noe lovforbud mot grøfting og oppdyrking av myr i Norge. Det norske myrselskap ble stiftet i 1902 for å fremme utnyttelsen av Norges myr- og torvressurser gjennom produksjon av brenntorv, torvstrø og myrdyrking. Grøfting av myr og myrskog for skogreisning og økt skogproduksjon fikk etter hvert et stort omfang, særlig etter 1930, og har siden pågått fram mot 2000-tallet gjennom offentlige tilskuddsordninger. Totalt er det laget ca. 165 000 km med myrgrøfter i Norge, de fleste før andre verdenskrig. Eksisterende lovverk forbyr nygrøfting av myr og sumpskog til skogbruksformål. Effekter av myrødeleggelseAlle inngrep på myr som medfører senkning av gunnvannsspeilet, er en trussel mot torvas evne til å lagre karbon. Ved grøfting og senkning av grunnvannspeilet endres og ødelegges plantedekket, og den underliggende torva oksideres og brytes ned til CO2 i løpet av få år, og slippes ut i atmosfæren som klimagass. Nydyrking av myr gir et totalt karbontap på 0,8 tonn pr. dekar og år de første årene (Grønlund et al. 2010). På verdensbasis forårsaker myrgrøfting nesten 10 % av alt CO2-utslipp i atmosfæren. Dyrket myr er også en kilde til betydelig utslipp av lystgass (N2O). Dette kompenseres i noen grad gjennom at utslippet av metan (CH4) reduseres sterkt etter drenering med senking av grunnvannsstanden og tilførsel av luftas oksygen. Netto klimagassutslipp ved dyrking av myr kan derfor grovt anslås til å ligge på ca. 2 tonn CO2-ekvivalenter pr. dekar og år (Grønlund et al. 2010). Uttak av torv til ulike formål (veksttorv, torvstrø, energiformål) har større negativ effekt. Samtidig vil oppdyrking av myr eller fjerning av torva hindre ny karbonlagring gjennom et levende mosedekke. Myrenes vannlagringskapasitet vil også reduseres betraktelig ved grøfting og drenering, og gradvis opphøre ved fjerning av torv. Opphøret av denne svampeffekten kan ha negativ effekt for regulering av flomsituasjoner. En mangler oversikt over de negative virkningene som myrødeleggelse har hatt på det biologiske mangfoldet av torvmoser i Norge. Men grøfting og drenering av skogsmyr på Østlandet er en opplagt årsak til at den rødlistede huldretorvmosen Sphagnum wulfianum der er forsvunnet fra mange av sine tidligere kjente voksesteder (Flatberg et al. 2010). Restaurering og revegetering av ødelagt myrArbeidet med restaurering og revegetering av ødelagte myrområder har som hovedformål å føre disse mest mulig tilbake til opprinnelig tilstand, dvs. gjenskape myrøkosystemet og biodiversiteten, inklusive kapasiteten til å lagre karbon. De primære tiltakene så langt har vært reetablering av mest mulig gunstige grunnvannsforhold, for eksempel gjennom avstenging av grøfter og andre kunstige dreneringsbaner, og revegetering gjennom flekkvis flytting av torvmoser fra egnede donorsteder med intakte mosematter/-puter. Dette blir gjort ved at det ca.10 cm øverste laget av levende torvmose blir fjernet flekkvis fra et voksested og overført til restaureringsområdet. I praksis blir dette utført slik at en torvmosematte/-pute tilsvarende ca. 1 m2 blir fragmentert i skuddbiter og spredd på 10–15 m2 bar torv, for eksempel ved bruk av en gjødselspreder. Forsøk har vist at et tynt dekke av halm virker gunstig på revegeteringen av torvmosene. Kanada, med store ødelagte myrområder, har vært en pioner i arbeidet med restaurering og revegetering av myr (Quinty & Rochefort 2003). Revegetering på denne måten har gitt myrstrukturer og myrtopografi som er sammenlignbar med naturlig myr, i løpet av få år (Pouliot et al. 2011b). Til sammenligning vil ødelagt myr overlatt til seg selv bruke mer enn hundre år på å revegetere. Men også i nordisk sammenheng er det utført vellykket restaurering av myr med torvmoseetablering (Haapalehto et al. 2011, Hedberg et al. 2012). På De britiske øyer har en også arbeidet med å finne fram til praktiske metoder for revegetering av store ødelagte myrområder med bruk av transplanterte torvmoser (Hinde et al. 2010). Forsøk med oppformering av utvalgte torvmoser på laboratoriebasis og innkapsling av store mengder av små skuddfragmenter i små, gelatinøse, næringskapsler (mikropropaguler) er en del av det forsøksarbeidet som pågår. Målet er at mikropropaguler skal kunne sprøytes ut over større ødelagte myrområder og gi grunnlag for ny etablering og vekst av torvmoser." Grunnvannets betydning for dannelse av torv, myr og torvmassivenheterSkal vi forstå hvordan torvmassiver dannes og overflateformene oppstår, må vi starte med grunnvannets rolle og forutsetningene for torvdannelse. Grunnvannsspeilet på myr, er øverste nivå der alle porer er vannfylte. Under grunnvannsspeilet er det sammenhengende vannsøyle. Men det betyr ikke at denne sammenhengende vannsøyla er homogen. Den kan godt være lagdelt, med ombrogent vann (nedbørsvann) øverst og minerogent vann (jordvann) under.
Akkumulering av organiske jordarter (torv) skjer når produksjonen av organisk stoff er større enn nedbrytingen. Etter dannelsesmåten deles myrene i gjenvoksningsmyr og forsumpningsmyr. 1. Gjenvoksningsmyrene. Disse har begynt som organiske avsetninger i vatn. 2. Forsumpningsmyrene. Der har torvdannelsen funnet sted på fastmark. Størstedelen av myrarealet i Norge er dannet på denne måten. 1. Gjenvoksningsmyrer:
2. Forsumpningsmyrer:
Myr og torv i Natur i NorgeLes mer om myr og torv fra en rapport i 2023 her:
https://ntnuopen.ntnu.no/ntnu-xmlui/handle/11250/3063175 "Myr defineres som et landområde med vegetasjon som krever fuktighet, og som danner torv. Torv er sted-egent akkumulert materiale, hvis tørrvekt utgjøres av mer enn 30 % dødt organisk materiale. Det er ingen krav til tykkelse på torvlaget for at noe skal kalles myr. Torvmark defineres som et landområde med eller uten vegetasjon, med et naturlig akkumulert torvlag på toppen. All myr er derfor torvmark, men torvmark trenger ikke være myr." […] "Natur i Norge versjon 3 (NiN 3) har to typesystemer for myr. Typesystemet for myrøkosystemer skiller mellom hovedtyper og grunntyper på bakgrunn av gradienter i vegetasjon og miljøforhold. Typesystemet for torv-massiv (myrlandformer) inneholder torvmassivenheter som er definert på grunnlag av forskjeller i hydrologi, hydromorfologi og hydrogenetikk. Et torvmassiv er en hydromorfologisk enhet med hydrologi og overflate-former som gjenspeiler samspillet mellom topografi, vanntilførsel og torvdannende prosesser." […] "Skillet mellom minerogen og ombrogen markvæte er fundamentalt, og er derfor lagt til grunn for å skille mellom fire grupper minerogene torvmassiv på den ene siden og to grupper ombrogene torvmassiv på den andre siden." Oversikt over torvmassivenheter (tidligere hydromorfologiske myrtyper)Skjematisk framstilling av 22 torvmassivenheter som inkluderes i NiN 3. Til venstre er torvmassivene vist i profil, og til høyre er viktige overflatestrukturer vist. Under de enkelte illustrasjonene er det oppgitt vanlig størrelse på torvmassivenhetene, og til venstre er vanlig torvdybde angitt (høydeskalaen er overdrevet). Lys brun = ombrogen torv; mørk brun = minerogen torv; grå = fastmark (mineraljord og berggrunn); blå = åpent vann; lys blå = is; grønn = trær.
Torvmarksmassiv i NiN"Torvmarksmassiv brukes for å beskrive myr som landform, og beskriver torvforekomstens egenskaper, ikke myrøkosystemet på torvoverflata. Typeinndelingen i Torvmarksmassiv er resultatet av de økologiske forholdene som har virket på myrdanning og myrvekst over tid."
Les om myr, torv og torvmassiver i ny bok fra Universitetsforlaget, utgitt 2024. Klikk på bildet, eller her, og les om boka, bestille den, og se et sammendrag og innholdsfortegnelse. Utdrag fra boka:Del 2 Naturvariasjon systematisert gjennom NiNØkoegenskaper(Side 72) "Den tredje hovedgrenen i naturmangfoldtreet, økoegenskaper, omfatter naturegenskaper som er resultatet av at abiotiske og biotiske egenskaper virker sammen. Begrepet økoklin brukes om samvariasjonen mellom en gradient i artssammensetning og en kompleks miljøgradient. Økoklinbegrepet er helt sentralt når vi skal sette naturegenskapene i system fordi det er nettopp dette som fremheves i naturtypedefinisjonen. Økoklinen er økoegenskapenes parallell til den abiotiske miljøvariasjonen og den biotiske artssammensetningsvariasjonen. Økokliner kan være menneskebetingete eller naturgitte, de kan deles inn i regionale, lokale og korttidsøkokliner. Det ligger i selve økoklinbegrepet at det i utgangspunktet er begrenset til gradvis (klinal) variasjon, men det brukes i en videre betydning som også inkluderer naturlig klassedelt variasjon. For å identifisere økokliner må vi vite hvilke arter som finnes innenfor et representativt utvalg av observasjonssteder i et område, og vi må kjenne miljøforholdene på disse stedene. Deretter må vi finne ut hvordan artssammensetningen og miljøforholdene samvarierer. Vi kan foreta en ordinasjonsanalyse av en matrise med artsmengdedata. Miljøvariabeldata som er samlet inn på de samme observasjonsstedene der artssammensetningen er registrert, kan brukes til å 'tolke' sammenhengen mellom gradienter i artssammensetning og de tilhørende miljøvariablene. Denne tolkete sammenhengen er en økoklin. Et eksempel på en økoklin er variasjonen i artssammensetning langs den komplekse miljøvariabelen 'kalkinnhold' i en skogsmark." […] Del 3 Landformvariasjon og marine vannmasserTorvmassiver(Side 145–159) "I NiN-systemet typeinndeles myrer både som landformer og som økosystemer (se kapittelet om våtmark i del 4). Myrer skiller seg fra de andre landformkategoriene ved at de nesten utelukkende består av torv, altså lite nedbrutte rester av planter som har vokst på stedet. Begrepet torvmassiv brukes i NiN 3 om myr som landform for å presisere at vi ser på torvforekomstens egenskaper og ikke myrøkosystemet på torvoverflata. Torvmassiver karakteriseres av overflateform, hydrologiske egenskaper og de prosessene som førte til dannelse av torv. Begrepet hydromorfologi brukes om torvmarkas overflateform (morfologi), som nesten alltid bestemmes av vanntilgangen (hydrologien). Skal vi forstå hvordan torvmassiver dannes og overflateformene oppstår, må vi starte med grunnvannets rolle og forutsetningene for torvdannelse. Grunnvann Torv I dagligtalen, og til dels også i NiN, brukes begrepene myr og torvmark om hverandre. Det er likevel en viktig nyanseforskjell. Torvmark er det tykke laget med akkumulert torv, mens myr er et område med våt torv der det akkumuleres torv i overflatelaget. Med unntak for ødelagte myrer har de aller fleste torvmarker torvproduserende vegetasjon på toppen og er derfor også myrer. Myrer (intakte torvmarker) kjennetegnes av kort avstand fra overflata ned til grunnvannsspeilet. Stikker vi et rør ned i en myr, vil det i løpet av noen timer etablere seg et vannspeil noen centimeter under torvoverflata. Dette er grunnvannsspeilet i myra, og under dette nivået er alle hulrom mettet med vann. Dannelse av torv Hvert torvmoseskudd består av en stengel med knipper av grener. I toppen av skuddet sitter knippene svært tett og danner et flatt eller hvelvet 'hode'. Under hodet sitter et vekstpunkt der stengelen forlenges, og det dannes nye grenknipper. Størstedelen av de små bladene på grenene utgjøres av døde, fargeløse celler, såkalte hyalinceller. Hyalincellene kan lagre vann, men vann lagres også i små hulrom mellom grenene og mellom tettstilte torvmoseskudd. Torvmoser kan derfor holde på en vannmengde som er inntil tjue ganger mosens tørrvekt. Nedbrytningen av plantemateriale på myra hemmes av at torvmosene forsurer miljøet når de tar opp Ca2+ og Mg2+ fra vannet ved ionebytte med H+. Torvmosenes årlige lengdevekst bestemmes av voksestedforholdene og kan bli så mye som 40 mm per år i en lavlandsmyr. Dette er imidlertid bare én blant mange faktorer som bestemmer torv-markas høydetilvekst. Allerede noen få centimeter under torvoverflata begynner nedbrytningen av visne torvmoser og annet dødt organisk materiale. Nedbrytningen fortsetter så lenge torvlaget er gjennomluftet, men er minimal under laveste grunnvannsspeilnivå. Derfor går det et viktig skille mellom det øverste, periodevis gjennomluftete torvlaget, akrotelmen, og det permanent vannmettete torvlaget, katotelmen. Mosematerialet blir dessuten stadig presset sammen under den økende tyngden av nytt materiale. Dersom vi ser bort fra nedbrytningen av materiale i katotelmen, bestemmes torvmassivets høydetilvekst av tilfanget av nytt organisk materiale til katotelmen. Sannsynligvis varierer torvtilveksten fra 0 mm til noe over 1 mm per år. Nedbrytningshastigheten for torv i katotelmen er anslått til mellom 1/1000 og 1/100 000 per år. Dersom torvtilveksten er 1 mm og nedbrytningshastigheten 1/10 000 per år, vil det innstille seg en likevekt mellom torvtilvekst og nedbrytning når torvlaget har blitt 10 m (10 000 mm) tykt. Myrer med så tykt torvlag finnes ikke i Norge. Typiske torvdybder i norske myrer er 2–5 m. Dette indikerer at norske myrer fremdeles akkumulerer torv og er i utvikling, og at torvtilveksten de fleste steder er mindre enn 1 mm per år. Torvdannelse fremmes av fuktig klima og en topografi som gjør at grunnvannsspeilet hele tiden ligger nær markoverflata. Sørlige og østlige deler av Europa mangler myr fordi klimaet er for varmt eller tørt. I tropiske strøk finnes myrer bare høyt over havet. I det boreale barskogsbeltet er klimaet de aller fleste steder fuktig og kjølig nok for torvdannelse, men produksjonen avtar med økende høyde over havet, og omkring grensa mellom lavalpin og mellomalpin bioklimatisk sone brytes plantematerialet ned like raskt som nytt blir produsert. Mellomalpin sone på fastlandet og mellomarktisk sone på Svalbard mangler derfor torvmark. Dannelse av torvmassiver Utviklingen av torvmassiver kan følge to prinsipielt forskjellige forløp: gjenvoksning (terrestrialisering) av et innsjøbasseng eller forsumping av tidligere fastmark (paludifisering). Terrestrialiseringsprosessen starter med at torvmoser etablerer seg langs kanten av et tjern. Det forutsetter at påvirkningen fra bølger eller is ikke er så sterk at torvmosemattene blir ødelagt av fysisk forstyrrelse. Gjenvoksning av tjern skjer derfor hovedsakelig langs små tjern som ligger godt beskyttet mot vind. Etter hvert som flytematter brer seg utover vannflata, transformeres tjernet til myr. Når bare et lite 'øye' av vann er igjen, har myra fått et karakteristisk utseende. Etter at øyet har lukket seg, fortsetter torva i de sentrale delene av gjenvoksningsmyra å vokse i høyden. Etter noen tusen år har myra fått en hvelvet torvkuppel som hever seg over vannspeilet i det som engang var et tjern, og til slutt hever torvkuppelen seg også over omgivelsene. Dersom vi stikker et rør ned i torva på toppen av kuppelen, vil vi se at det fortsatt er kort vei ned til grunnvannsspeilet. Det viser at det har etablert seg et konvekst grunnvannsspeil i den hvelvete myrkuppelen. Vannspeilet kan heve seg over omgivelsene fordi torva er så kompakt at den hindrer sideveis vannbevegelse. Nedbør som faller på den konvekse myroverflata, renner derfor ikke av, men bidrar til å opprettholde det høye grunnvannsnivået. I dette stadiet får det øvre torvlaget kun tilført nedbørvann. Når torvlaget har blitt så tykt at overflata hever seg klart over fastmarka omkring, mister grunnvannet i overflatetorva kontakten med jordvannslaget, og vi har fått en nedbørsmyr. Alle torvmarker får tilført ombrogent vann, men nedbørsmyrer tilføres bare vann med nedbøren. Forsumping på fastmark forutsetter enten at grunnvannsspeilet ligger såpass nær markoverflata at betingelsene for dannelse av våtmark blir oppfylt, eller at klimaet er fuktig nok til å opprettholde en våtmark uten tilførsel av grunnvann. Forsumping på tidligere fastmark i flatt terreng, det vil si med topogen markfukting, skjer over hele landet uavhengig av klimaforholdene. Forsumping i hellende terreng forutsetter at klimaet er fuktig nok til å opprettholde en permanent fuktig mark også når topografien gjør at vannet renner vekk. Jo sterkere helning, desto fuktigere klima kreves for å sette i gang torvdannelse. Langs vestkysten av Norge fra Ryfylke til Helgeland er klimaet så fuktig at også bratte skråninger kan forsumpes. Klimaendringer er den viktigste årsaken til begynnende forsumping av fastmark. Overgangen til et litt kjøligere og fuktigere klima for ca. 2500 år siden førte til økt forsumping over hele landet. Også lokale forhold kan initiere forsumping, for eksempel stormfelling. Myrer som er dannet ved forsumping av tidligere fastmark, både i flatt og i hellende terreng, kan utvikle seg til nedbørsmyrer hvis torvlaget blir tykt nok til at overflatetorva mister jordvannskontakten og tilførselen av jordvann forhindres. Myrenes hydromorfologi NiN skiller mellom fire ulike mikrostrukturer: LO-MY Mykmatte, LO-FA Fastmatte, LO-TU Tue og LO-LB Løsbunn. Mykmattene har løs torv som ofte oversvømmes og kan ikke bære et menneske. Fotspor i en mykmatte er derfor synlige i mange måneder. Fastmattene har fast, gjennomluftet torv som bærer et menneske slik at fotspor raskt viskes ut. Tuene hever seg over fastmattene og har kompakt torv. Løsbunn er en samlebetegnelse på områder med naken torv og kan finnes blant mykmatter og fastmatter. Myrstrukturer utgjør karakteristiske samlinger av mikrostrukturer. Eksempler på myrstrukturer er LO-SF Fastmattestreng, LO-ST Tuestreng, LO-HØ Hølje og LO-FL Flark. Strenger er lange, sammenhengende fastmatter eller tuer som er atskilt av våtere forsenkninger. De våte forsenkningene kalles høljer på nedbørsmyr og flarker på jordvannsmyr. Både høljer og flarker kan inneholde gjøler, det vil si små dammer med torvbunn. Fordelingen av myrstrukturer er ikke tilfeldig. Figurene 3.37 og 3.38 viser langstrakte strukturer på tvers av myroverflatas helningsretning. Utviklingen av strukturer starter et stykke ut i myrdannelsesprosessen. Når det regner mye, renner overskuddet av vann nedover myroverflata dersom det er mulig. Men torv er et lite vanngjennomtrengelig substrat, og vannet slipper bare gjennom de aller øverste centimeterne der torva er løs. Når overflatevann møter ugjennomtrengelig torv, spres vannet sideveis på tvers av myroverflatas helningsretning. Dersom torvmosematta blir stående under vann i lengre tid, hemmes mosenes vekst. Torvmosetilveksten reduseres ytterligere fordi torvmoser i og langs kanten av islagte gjøler utsettes for isskuring og andre former for frostbetinget forstyrrelse. De vannfylte forsenkningene fortsetter derfor å utvide seg sideveis. Dersom vannansamlingene på myroverflata blir permanente gjøler, forsterkes prosessen, og torva eroderes både fra kantene og fra bunnen. Gjølene vokser seg dermed større og dypere inntil det eventuelt inntreffer en balanse mellom torvtilveksten i strengene og erosjonen i gjølene. Begrepet myrsegment brukes om områder med ensartet fordeling av myrstrukturer. Hvert av de to avgrensete områdene i figur 3.38 utgjør et myrsegment. Eksempler på myrsegmenter er den tørrere, tuedominerte kantskogen (LO-KA) med spredte trær og laggen (LO-LA) med jordvannsmyr som skiller nedbørsmyr fra fastmark. Sammen med en åpen myrflate utgjør kantskogen og laggen en hydromorfologisk enhet på det fjerde skalanivået: torvmassiv. Torvmassiver er klart avgrensete, hydromorfologisk definerte landformer som består av myrsegmenter som har felles vannhusholdning. Torvmassivene er viktige forvaltningsenheter fordi påvirkning av én del av et torvmassiv også påvirker andre deler, for eksempel ved grøfting. Begrepet myrkompleks brukes om et geografisk avgrenset område med sammenhengende torvmark. Typeinndeling av torvmassiver NiN skiller mellom åtte torvmassivgrupper på grunnlag av dominerende vanntilførsel og viktigste torvmarksdannende prosesser. Overveiende minerogene torvmassiver omfatter fire grupper, og torvmassiver dominert av ombrogen vanntilførsel omfatter to grupper. De to siste gruppene består av torvmassiver med betydelig innslag av både minerogen og ombrogen torv. Til sammen inneholder typesystemet for torvmassiver 22 torvmassivenheter. Minerogene torvmassiver Som vi har forklart tidligere, starter all torvdannelse med minerogen torv. Alle torvmassivenheter har derfor utviklet seg fra torvmassiver i denne gruppa. Gruppe TM-A Topogene torvmassiv har flatt grunnvannsspeil. Denne gruppa inkluderer TM-A01 Gjenvoksningsmyr, som er et resultat av at et tjern fylles med torv, og TM-A02 Flatmyr, som er et resultat av forsumping på tidligere fastmark med flatt grunnvannsspeil, for eksempel i svake forsenkninger i terrenget. I fuktig klima kan som sagt også skråninger forsumpes. Resultatet blir TM-B Soligene torvmassiv. Størstedelen av Norge har et klima som er fuktig nok til at det kan dannes myrer i hellende terreng på steder med grunnvannsspeil nær markoverflata. Torvmassivenheten TM-B03 Bakkemyr forutsetter en gjennomsnittlig helning > 3°, mens myrer med bevegelig grunnvann og helning 0–3° betegnes TM-B01 Slakmyr. Selv om overgangen mellom flatmyr og slakmyr og videre til bakkemyr er helt gradvis, er det en viktig hydrologisk forskjell mellom topogene og soligene torvmassiver fordi bevegelig vann har andre egenskaper enn stagnerende vann. I nedbørrike, høyereliggende områder med slake terrengformer, for eksempel i indre deler av Trøndelag, kan bakkemyrer dekke størstedelen av landskapet. Bakkemyrene har ofte relativt grunn torv fordi vannet som renner gjennom myra, er mer oksygenrikt og rikere på mineralnæring enn stagnerende vann, noe som fremmer nedbrytning. I de klimatisk fuktigste delene av Norge, det såkalte midtfjordsbeltet innenfor kysten, er det ikke uvanlig å finne små, smale bakkemyrer med helning på 30° eller mer. Bakkemyrer med regelmessig ordnete strenger og flarker fordeles på enhetene TM-B04 Flarkmyr og TM-B05 Strengmyr. Begge disse finnes i hele området der det finnes bakkemyrer, men er vanligst i Midt- og Nord-Norge og i litt høyereliggende strøk på Østlandet. Gruppa TM-C Kildetorvmassiv omfatter bare én enhet, TM-C01 Torvkilde, med tykk mosematte som helt eller delvis dekker vannspeilet (figur 3.40). Torvkilder finnes over hele landet, men er vanligst i nordboreal og nedre del av lavalpin bioklimatisk sone. Små torvmassiver kan også dannes på steder som regelmessig påvirkes av flomvann fra innsjøer og elver eller tilføres tidevann. Slike torvmassiver utgjør gruppa TM-D Flommyrtorvmassiv, som omfatter henholdsvis TM-D01 Innsjøflommyr, TM-D02 Elveflommyr og TM-D03 Saltflommyr. Innsjøflommyr kan være et tidlig stadium i utviklingen av en gjenvoksningsmyr. Elveflommyr har torv med høyt innhold av mineralkorn (silt, sand og grus) som transporteres med elvevannet og fanges mellom torvmoseskuddene i flomperioder. Saltflommyrer er nokså sjeldne, men finnes på beskyttete steder i øvre del av fjærebeltet, mest i Nord-Norge. Ombrogene torvmassiver I et sent stadium i utviklingen fra minerogen til ombrogen myr, for eksempel fra gjenvoksningsmyr via flatmyr, kan et karakteristisk myrmassiv med hvelvet, ombrogen overflate (et såkalt høgmyrmassiv) begynne å ta form. TM-H Høgmyrmassiv er én av de to gruppene av ombrogene torvmassiver. De fleste høgmyrmassiver har en lagg (LO-LA) mot fastmarka. Laggen har minerogen torv og tar imot jordvann fra fastmarka og eventuell overflateavrenning fra det sentrale nedbørsmyrområdet. Den ombrogene myrflata kan være tuedominert eller ha store mykmatter med gjøler på steder som mottar mye vann, eller der videre avrenning hindres. Høgmyrmassiver omfatter betydelig hydromorfologisk variasjon og fordeles på seks enheter med karakteristisk morfologi (se figur 3.39). TM-H01 Atlantisk høgmyr (se figur 3.38) mangler både kantskog og lagg og omfatter svakt hvelvete myrer med stor variasjon i fordelingen av strukturer på overflata. Grensa mot tilgrensende torvmassiver er ofte uskarp, og det kan være vanskelig å vite hvor jordvannstilførselen stopper. TM-H04 Konsentrisk høgmyr har strukturer som er tydelig organisert i konsentriske sirkler omkring det høyeste punktet, mens TM-H01 Atlantisk høgmyr og TM-H03 Platåhøgmyr har en mer plan, åpen myrflate i sentrum og mindre regelmessig orientering av strukturene. Konsentrisk høgmyr og platåhøgmyr har ofte kantskog (LO-KA), i hvert fall langs deler av kanten mot laggen. Atlantisk høgmyr forekommer i kyststrøk fra Agder til Andøy lengst nord i Nordland. Konsentrisk høgmyr er mer eller mindre begrenset til Østlandet, mens platåhøgmyr også finnes i Midt-Norge. TM-H05 Skogshøgmyr omfatter hvelvete nedbørsmyrer som i sin helhet er dekket av en skrinn, sentvoksende furuskog. Denne typen torvmassiv forutsetter at overflatetorva i perioder blir tørr nok til at furutrærne kan vokse seg så høye og stå så tett over det meste av myra at det dannes et skogsmarksøkosystem. I tillegg må de klimatiske betingelsene for torvdannelse være oppfylt. De få forekomstene av skogshøgmyr i Norge finnes i lavlandet Østafjells, særlig i indre Østfold. De utgjør ytterpunktet i et utbredelsesområde som strekker seg sør- og østover mot Sør-Sverige og Baltikum. Høgmyr kan også utvikles i hellende terreng, oftest med slakmyr som utgangspunkt. Da dannes TM-H06 Eksentrisk høgmyr, som kjennetegnes av lineære eller buete, parallelle strukturer på den åpne myrflata som heller fra et høyeste punkt nær kanten. Eksentriske høgmyrer forutsetter forsumping i hellende terreng og er dermed knyttet til fuktigere klima enn konsentriske høgmyrer og platåhøgmyrer. De forekommer ofte over svake rygger i terrenget, skjermet mot tilførsel av vann fra fastmarka, og finnes først og fremst i sørboreal bioklimatisk sone på Østlandet og Midt-Norge. Myrkompleks med eksentriske høgmyrer på ryggene og slakmyrdråg i skålformete forsenkninger er ikke uvanlig i småkuperte åslandskaper. TM-H02 Kanthøgmyr er en spesiell type høgmyrmassiv som bare finnes spredt langs kysten. I midtfjordsbeltet langs vestkysten av Norge regner det så mye at nesten all mark kan forsumpes. Der kan det til og med dannes torvmark direkte på nakent fjell. Dersom de gunstige forholdene for forsumping fortsetter i lang tid, vil en stadig en større del av landskapet forsumpes. Etter hvert flyter torvmarksflekkene sammen til et lappeteppe av torvmark og fastmark, og til slutt kan hele landskapet være dekket av torvmark. Avhengig av topografi, klima og andre forhold vil denne torvmarka kunne vokse i høyden og over tid utvikle seg til en myr som dekker store områder. TM-G01 Terrengdekkende myr omfatter sammenhengende torvmassiver som følger terrengets form og er dominert av nedbørsmyrtorv, men har jordvannsmyrpartier i forsenkningene. Terrengdekkende myr er eneste enhet i torvmassiv-gruppa TM-G Ombrogene, ikke hvelvete (terrengdekkende) torvmassiv. Torvmassiver med både minerogen og ombrogen torv Den siste torvmassivgruppa, TM-F Mellomstillingsmassiv, omfatter én enhet, TM-F01 Mellomstillingsmyr. Dette er torvmassiver med både minerogene og ombrogene deler. Enheten omfatter umodne systemer som er på vei til å bli høgmyr. Noen av dem vil kanskje fortsette å inneholde minerogene og ombrogene flekker i en kompleks mosaikk, mens andre vil utvikle seg videre til en høgmyr. Planmyr har vært brukt som betegnelse for mellomstillingsmassiver med tilnærmet plan overflate. De har gjerne minerogene partier nær kanten og utydelig avgrensete ombrogene partier i de sentrale delene. Slike myrer er ofte preget av erosjon og naken torv på steder med stor avrenning. Alle mulige overganger mot terrengdekkende myr og høgmyr finnes. I internasjonal myrlitteratur brukes begrepene bog og fen om henholdsvis ombrogene og minerogene torvmassiver. I Sverige finnes tilsvarende folkelige begreper, henholdsvis mosse og kärr, mens norsk mangler presise, tilsvarende begreper." […] Del 4 Økologisk mangfoldDe viktigste komplekse miljøvariablene[…] (Side 171–172) "For å kunne opprettholde sine livsfunksjoner må alle organismer få oppfylt noen helt grunnleggende betingelser. Den aller viktigste er sannsynligvis behovet for næring. Alle organismer forholder seg dessuten til vann som essensiell ressurs eller som oppholdssted. Akvatiske arter er tilpasset et liv i vann, mens for de fleste terrestriske arter er vann en ressurs som er viktig for både vekst og reproduksjon. En god del arter er dessuten amfibiske, det vil si at de er tilpasset et liv i overgangen mellom vann- og landøkosystemer. Siden vann og næring bestemmer de fysiologiske toleransegrensene for veldig mange organismer, gir dette stor variasjon i artssammensetningen langs de komplekse miljøvariablene som representerer disse egenskapene: fra tørre til våte steder eller fra steder med lite til mye næring. Vi finner for eksempel orkidéen myrflangre Epipactis palustris på kalkrik våtmark, men ikke på myr som er næringsfattig, eller i skogen der det er tørt. Det skyldes at denne arten er avhengig av fuktighet, høy pH og god næringstilgang, miljøforhold som i NiN representeres av miljøvariablene vannmetning og kalkinnhold. Flere miljøvariabler er relevante for mange økosystemer. Det er derfor nyttig å gi en felles beskrivelse av de viktigste miljøvariablene før vi ser nærmere på hvordan de påvirker artssammensetningen i de enkelte økosystemene. Vi har gruppert disse komplekse miljøvariablene i fem temaer: hvor stor del av tiden bakken er dekket av vann, vanntilførsel til landsystemer, næringsstofftilførsel, saltinnhold og dominerende kornstørrelse (størrelse på partikler som stein og liknende). I dette kapittelet skal vi se på hvordan variasjonen i disse temaene beskrives i NiN, og hvordan temaene kobles til hvert sitt sett av komplekse miljøvariabler. På denne måten forklarer vi begrepene som blir tatt i bruk i de påfølgende kapitlene, først for å skille mellom natursystemhovedtypegrupper og deretter for å skille mellom hovedtyper og grunntyper på de neste nivåene." […] (Side 175) "Gradienten fra mykmatte til tue i myr En tydelig forskjell fra tidevanns- og flombeltene er at variasjonen i vannstand gjennom året er mye mindre, helt ned til 10 cm. Det er oftest liten horisontal vannbevegelse på en myr; endringer i grunnvannsnivået avhenger derfor av tørke eller vann som tilføres gjennom nedbør og snøsmelting. Forskjellen mellom høyeste og laveste avstand til grunnvannsspeilet er noe større under torvmosedominert og tørrere vegetasjon siden torva består av fast materiale med porer som kan fylles med vann. Aller størst er forskjellen under tuene, der den kan bli 30 cm eller mer. Mykmatter med sammenhengende dekke av torvmoser finnes vanligvis ikke på steder som er oversvømt mer enn halve tiden. Det skyldes at is- og vannerosjon kan ødelegge mattestrukturen. Omtrentlig nedre grense for mykmatter sammenfaller derfor stort sett med grensa mellom hydrolitoral og geolitoral (se figur 4.1). Grensa mellom fastmatte og tue er ofte tydelig markert ved den nedre grensa for vital røsslyng Calluna vulgaris, som tilsvarer middelflomnivået. Det er bare unntaksvis og i veldig korte perioder at vannstanden når opp til tuenivået. Mykmattenivå, nedre og øvre fastmattenivå tilsvarer dermed de tre geolitorale beltene i tidevanns- og flombeltet med hensyn til oversvømmelsesvarighet. På myr er disse beltene veldig distinkte til tross for at de er svært komprimert, og hvert av dem typisk har en vertikalutstrekning på bare 2–4 cm. Dette viser at forskjeller i varigheten av oversvømming er like viktig på myr som i tidevanns- og flombeltene. Tuene oversvømmes ikke, men har likevel en vertikalvariasjon i artssammensetning som gjenspeiler variasjon i fuktighetsforholdene." […] Vanntilførsel til landsystemer(Side 177) Vanntilførsel til jorddekt fastmark […] (Side 190) Grensa mellom fastmarkssystemer og våtmarkssystemer"NiNs definisjon av NA-V Våtmarkssystemer krever at organismene har god og stabil vanntilgang og er tilpasset et liv under vannmettete forhold. Dette finner vi på steder der grunnvannsspeilet er tilstrekkelig nær markoverflata eller har god tilførsel av overflatevann. Våtmarkssystemer mangler imidlertid vannlevende organismer og har vanligvis en vanndekning mindre enn 50 % av tiden. Våtmarksgrensa trekkes mellom to trinn langs variabelen LM-VM Vannmetning (se tabell 4.1); LM-VM_c fuktig (mark) tilhører fastmarkssystemer, og overgangstrinnet LM-VM_z våt (mark) tilhører våtmarkssystemer. Hovedtypegruppa våtmarkssystemer inkluderer all myr, det vil si torvmark der det akkumuleres torv. Men – all torvmark er ikke våtmark. Under spesielle forhold kan nemlig tykke organiske jordlag som tilfredsstiller definisjonen av torv, også dannes på fastmark (se kapittelet om torv i del 3). Hovedtypegruppe NA-V inkluderer også kilder, det vil si mark med LM-KI Kildevannspåvirkning som minst er LM-KI_d klar." […] Natursystem – våtmarkssystemer(Side 348–362) "Myrer kan fremstå som våte, ugjestmilde og utilgjengelige. Historier om hester og mennesker som har 'gått i myra', nasjonalromantiske malerier fra 1800-tallet og bilder av endeløse flater med små, krokete furutrær bygger opp under denne forestillingen. Turstier legges helst rundt myrer, og myrene klopplegges gjerne hvis det ikke er mulig å gå rundt dem. De fleste som ferdes i naturen, unngår derfor nærkontakt med myr som økosystem. Myr plasseres i hovedtypegruppa våtmarkssystemer (NA-V) i NiN. Dette er særpregete økosystemer med mange viktige økosystemfunksjoner. Vi har tidligere beskrevet myr fra et hydromorfologisk ståsted, det vil si som torvmassiv (se del 3). I dette kapittelet skal vi se nærmere på hva som kjennetegner våtmarksøkosystemer, og forklare den viktige rollen de spiller i naturen, i både en nasjonal og en global sammenheng. Våtmarkssystemer i et nasjonalt og globalt samfunnsperspektiv Grove anslag indikerer at 5–8 % av det globale landarealet består av våtmark i vid forstand. Omtrent 2/3 av dette finnes i tempererte, boreale og subarktiske klimasoner på den nordlige halvkule. Våtmarkssystemene med sine torvlagre inneholder trolig 25 % av den delen av jordas karbonlager på 2500 Gt (1 Gt = 109 tonn) som er bundet opp i jordsmonnet. Antakeligvis har mer enn halvparten av det opprinnelige europeiske myrarealet gått tapt på grunn av utnytting til ulike formål. Tilsvarende anslag for Norge er svært usikre, men et tall på 7000 km2 har blitt foreslått. Det totale norske myrarealet (inkludert grøftet og oppdyrket myr) beregnes til å være ca. 15 % eller mellom 45 000 og 50 000 km2 av et samlet landareal på 323 782 km2. Av dette er altså anslagsvis 1/7 utnyttet til ulike formål, mens det resterende, ca. 12 % av landarealet, er mer eller mindre intakt myr. Dette kan fordeles på jordvannsmyr (8,5 %) og nedbørsmyr (3,2 %). Ikke tresatt, åpen myr utgjør 8,8 % og myrskogsmark 2,9 %. Frem til siste halvdel av 1960-tallet ble myr først og fremst sett på som en ressurs vi kunne utnytte. Da skjedde et raskt fokusskifte fra utnyttelse til vern. I løpet av 1970- og 1980-tallet ble myrer vernet basert på representativitet og sjeldenhet. Frem mot vår tid har stadig mer kunnskap om myrøkosystemenes egenart og økologiske funksjon ført til en økt bevissthet om å ta vare på myr. De pågående globale klimaendringene har vært en viktig pådriver for dette fordi torvmarkenes funksjon som karbonlager spiller en nøkkelrolle i det globale karbonkretsløpet. Det lagres fortsatt karbon i norske torvmarker, noe som vil fortsette i tusener av år dersom denne funksjonen opprettholdes. På den annen side vil tining av permafrost og økt nedbrytning av torv i et varmere klima kunne gjøre at myrene går fra å være et sikkert karbonlager til å bli en kilde til store karbonutslipp. Nedbrytning av torv i miljø med lite oksygen har nemlig metan som sluttprodukt, en gass som har mye større drivhuseffekt enn karbondioksid. Myrene har også andre viktige funksjoner enn å ta karbon ut av kretsløpet. Den periodevis gjennomluftete overflatetorva fungerer nemlig som en svamp i terrenget. I perioder med mye nedbør fanger myrene opp store mengder vann som dermed slippes ut med forsinkelse til bekker og elver. Intakte myrer er derfor viktig for å redusere skadevirkningene av flomtopper, særlig dersom det blir mer ekstremvær fremover. Økt forståelse for myras viktige funksjoner har ført til strengere restriksjoner på ødeleggende bruk av torvmark, enten det gjelder utbygging av veier, industriområder, boligområder eller oppdyrking. Grøfting av myr for å anlegge skog ble forbudt i 2006. De siste årene har det blitt gjennomført flere restaureringstiltak på grøftet myr. Store maskiner brukes blant annet til å bygge demninger for å gjenopprette grunnvannsspeilet. Restaurering av myrøkosystemer krever derimot at torvproduksjonen igangsettes igjen. Bare fremtiden vil kunne gi svar på om dagens initiativer med hensyn til myrrestaurering lykkes. Det er svært vanskelig å gjenopprette myr som torvproduserende økosystem, noe som krever inngående kunnskap om den fininnstilte balansen mellom vanntilgang og torvproduserende vegetasjon. Normal variasjon i våtmarkssystemer De to lokale komplekse miljøvariablene LM-KA Kalkinnhold og LM-TV Tørrleggingsvarighet er spesielt viktige for artssammensetningen i våtmarkssystemer (se innledningskapittelet i del 4). Kalkinnhold uttrykker kompleks samvariasjon mellom pH i torv og vann, konsentrasjonen av kationer som Ca2+ og Mg2+, nitrogenmengde og innholdet av organisk materiale i torva. Variasjonen i kalkinnhold i våtmarkssystemer spenner over basistrinnene KA_a til KA_i, tilsvarende pH-intervallet fra ca. 4 til 8. Omtrent alle plantearter som vokser på myr, har klare preferanser med hensyn til kalkinnhold. Det tydeligste skiftet i artssammensetning skjer omkring midten av gradienten (KA_e); det blir mindre torvmoser Sphagnum spp, som erstattes av mattedannende bladmoser som stormakkmose Scorpidium scorpioides, rødmakkmose Scorpidium revolvens, myrstjernemose Campylium stellatum og gullmose Tomentypnum nitens. Rikmyrmosene kalles ofte brunmoser, som brukes som et samlebegrep for bladmoser som ikke tilhører torvmosene. Kalkrike myrer har ofte tynnere torvlag enn kalkfattige myrer fordi brunmosene produserer mindre torv enn torvmosene. Brunmoseskuddene er mindre, mattene er løsere og bedre gjennomluftet slik at nedbrytningen går raskere enn i tette torvmosematter. Grunn, kalkrik torv har større innhold av mineralmateriale enn kalkfattig torvmosetorv. Mens nitrogen begrenser produksjonen på kalkfattige myrer, er ofte fosfor begrensende faktor på svært kalkrike myrer fordi fosfor og kalsium danner tungtløselige salter. Mengden arter som potensielt kan finnes på et sted, øker med økende kalkinnhold. Antall arter som finnes på en liten flekk, for eksempel i en 1 m2-rute, blir imidlertid ikke nødvendigvis større. Begrepene fattigmyr og rikmyr har vært brukt i fennoskandisk myrlitteratur i over hundre år og viser til det økende artsantallet og ikke økende kalkinnhold som sådan. Det er imidlertid sterk samvariasjon mellom artstilfanget og kalkinnholdet. Fattigmyrer (LM-KA_abc) kjennetegnes av torvmosedominans og lite næringskrevende arter som flaskestarr Carex rostrata og sveltstarr C. pauciflora. Rikmyrer (LM- KA_ghi) kjennetegnes av en artsrik flora med orkidéer som engmarihånd Dactylorhiza incarnata og myrflangre Epipactis palustris i tillegg til mange mindre vanlige starrarter som for eksempel hårstarr C. capillaris, kjevlestarr C. diandra, taglstarr C. appropinquata og engstarr C. hostiana. LM-TV Tørrleggingsvarighet er den andre lokale komplekse miljøgradienten som er spesielt viktig for artssammensetningen i jordvannsmyr. Den påvirker artssammensetningen på mikroskala eller fin lokal skala. Tørrleggingsvarighet uttrykkes som medianavstanden fra torvoverflata til grunnvannsspeilet og tilsvarer varigheten av oversvømt torvoverflate. De tre mikrostrukturene LO-MY Mykmatte, LO-FA Fastmatte og LO-TU Tue gjenspeiler variasjonen langs denne gradienten. Artssammensetningen skiftes nesten fullstendig ut fra de våteste mykmattene som oversvømmes omtrent halve vekstsesongen (LM-TV_c), til de høyeste tuene (LM-TV_k) som kan rage 20 cm eller mer over høyeste grunnvannsstand. Små forskjeller i plassering relativt til grunnvannsspeilet gir dermed store utslag på artssammensetningen i myk- og fastmatter fordi disse oversvømmes i perioder. Forskjellen i avstand til grunnvannsspeilet er bare ca. 10 cm mellom nedre mykmatte og grensa mellom fastmatte og tue som markerer grensa for torv som oversvømmes i veldig våte perioder. I løpet av disse ti centimeterne skiftes mesteparten av artssammensetningen ut. En hårfin balanse mellom torvmoseartene i bunnsjiktet i fattigmyr og i nedbørsmyr bestemmer hvordan de fordeler seg langs denne gradienten. Torvmoser tåler både oversvømming og uttørring. Torvmosemattene endrer gradvis struktur. På mykmattenivå vokser vasstorvmose Sphagnum cuspidatum, lurvtorvmose S. majus eller bjørnetorvmose S. lindbergii i svært løse matter, ofte med liggende, spredte skudd. På fastmattenivå vokser vortetorvmose S. papillosum, rødtorvmose S. rubellum eller dvergtorvmose S. tenellum i fastere matter med opprette skudd. I tuer finner vi kompakte matter av rusttorvmose S. fuscum eller kysttorvmose S. austinii. Balansen mellom disse artene styres av avveiningen mellom veksthastighet og evnen til å tåle uttørring. Dette er to egenskaper som knapt kan optimaliseres på samme tid fordi arter som vokser raskt i løse matter, holder dårligere på vannet enn saktevoksende arter som vokser i kompakte tuer. De aller fleste moser er ektohydriske, det vil si at de tar opp vann og næringsstoffer gjennom overflata (se delkapittelet om vanntilførsel til landsystemer). Når de er fuktige, vokser mykmatteartene raskere enn tueartene, men mattestrukturen gjør at mykmattene og fastmatter dominert av den lille, løstvoksende arten dvergtorvmose Sphagnum tenellum tørker raskere ut enn øvrige fastmatter. Kompakte rusttorvmosedominerte tuer tørker ut sist på grunn av evnen til kapillært vannopptak fra dyp ned til 50 cm. Arter som vokser i løse matter som tørker raskt ut, har klare øvre grenser mot større tørrleggingsvarighet fordi de har dårligere evne til kapillært vannopptrekk. Torvmoseartenes nedre grenser mot mykmattene er derimot mindre skarpe. Det kan skyldes at tueartene tåler langvarig oversvømmelse dårlig, eller at arter med optimum nærmere grunnvannsspeilet vokser raskere og dermed har bedre konkurranseevne i våtere miljøer enn saktevoksende tuearter. Karplantenes grenser langs tørrleggingsvarighetgradienten bestemmes av miljøstress, det vil si hvor godt de tåler langvarig oversvømming av overflatetorva der artene har røttene sine. Den tydeligste utskiftingen av karplantearter skjer mellom fastmatte og tue, det vil si mellom torv som regelmessig oversvømmes, og torv som ikke oversvømmes mer enn eventuelt noen få dager i året. Mange karplanter, først og fremst vedplanter med mykorrhiza som krekling Empetrum nigrum og røsslyng Calluna vulgaris, mangler gjennomluftingsvev i røttene og tåler derfor dårlig langvarig oversvømmelse av rotsjiktet. Disse artene holder seg dermed til tuene. For at en furuplante skal vokse og bli et tre, kreves større, sammenhengende tuepartier. Andre arter uten gjennomluftingsvev, som bjønnskjegg Trichophorum cespitosum og molte Rubus chamaemorus, har litt større toleranse overfor oversvømming og finnes derfor også i øvre fastmattenivå. Mykmattene domineres av sumpplanter (helofytter) og andre arter som er tilpasset et liv med røttene i vann, for eksempel flaskestarr Carex rostrata, dystarr C. limosa, hvitmyrak Rhynchospora alba og sivblom Scheuchzeria palustris. Bare noen få arter, først og fremst torvull Eriophorum vaginatum og hvitlyng Andromeda polifolia, er mer eller mindre likegyldige overfor variasjon i tørrleggingsvarighet. Begge disse er imidlertid vanligere i fastmatter og tuer enn i mykmatter. Hovedtypen NA-VA01 Åpen jordvannsmyr (og NA-VC01 Åpen nedbørsmyr) inneholder også variasjon langs en tredje gradient i artssammensetning: fra myrkant til myrflate. I NiN kalles denne gradienten LM-MF Myrflatepreg. Begrepene myrflate og myrkant har vært brukt i skandinavisk myrlitteratur siden første halvdel av 1900-tallet og er godt innarbeidet. De blir derfor videreført i NiN til tross for at mange oppfatter begrepene som forvirrende. Områder med myrkantpreg kan nemlig finnes langt fra kanten av myra, og områder med myrflatepreg kan finnes nær kanten mot fastmark. Det vanligste er imidlertid at denne vegetasjonsgradienten også gjenspeiler plasseringen på myra. Myrkantpreget våtmark har mange felles arter med fuktig skogsmark, for eksempel myrfiol Viola palustris, skogstjerne Lysimachia europaea, tepperot Potentilla erecta, enghumleblom Geum rivale og tvaretorvmose Sphagnum russowii. Andre arter som karakteriserer myrkant, er også vanlige langs elver og innsjøer, for eksempel myrhatt Comarum palustre, sennegras Carex vesicaria og elvesnelle Equisetum fluviatile. Typiske myrflatearter er mer eller mindre begrenset til myr, for eksempel bjønnskjegg Trichophorum cespitosum, rome Narthecium ossifragum, dystarr Carex limosa og rødtorvmose Sphagnum rubellum. Få myrarter er likegyldige overfor denne gradienten som deles inn i fire basistrinn fra typisk myrkantpreg (MF_a) til typisk myrflatepreg (MF_d). De økologiske årsakene til variasjon langs denne artssammensetningsgradienten er fortsatt mangelfullt forstått og sannsynligvis et resultat av flere enkeltmiljøvariabler som i større eller mindre grad varierer sammen. Hovedtypen NA-VB01 Myr- og sumpskogsmark representerer et ytterpunkt langs myrflatepreg-gradienten; all skogsmark på torvmark er myrkantpreget. Innholdet av typiske arter for fastmarksskogsmark er klart større i myr- og sumpskogsmark enn i åpen jordvannsmyr med myrkantpreg. Vanlige trær i boreal skog, som gran Picea abies, furu Pinus sylvestris og bjørk Betula pubescens, mangler gjennomluftingsvev i røttene. Store, sammenhengende områder som ikke oversvømmes, er derfor en forutsetning for at trær på torvmark skal oppnå den høyden og tettheten som må til for å tilfredsstille definisjonen av skogsmark. Myr- og sumpskogsmark har dermed oftest en tørrleggingsvarighet tilsvarende tuenivå, mens sumpskog på mer kalkrik mark kan ha trær som står på høye, stylteformete tuer, for eksempel gran Picea abies og svartor Alnus glutinosa. Sannsynligvis er dette mulig fordi et stort rotsystem ikke er nødvendig når marka har høyere mineralnæringsinnhold. NA-VB01 Myr- og sumpskogsmark viser samme variasjon relatert til LM-KA Kalkinnhold og LM-TV Tørrleggingsvarighet som NA-VA01 Åpen jordvannsmyr, men med en sterk forskyvning mot dominans av tuenivå. Våtmarkssystemer er suboptimale voksesteder for de fleste treslag, kanskje med unntak for svartor. Aktivt voksende torvmosematter i næringsfattig miljø er nemlig et substrat der det er vanskelig for frøplanter å etablere seg. Dødeligheten er stor på grunn av faren for å bli overvokst av moser. Småplantene kan også bukke under på grunn av langvarig oversvømmelse. Furu finnes imidlertid ofte som spredte småbusker på åpne, kalkfattige myrflater under skoggrensa. På slike steder, der vekstvilkårene er helt marginale, kan furuer som er mindre enn én meter høye, likevel være mer enn 100 år gamle. Spesiell variasjon i våtmarkssystemer Nedbørsmyr Nedbørsmyrtorv har lavere konsentrasjon av mineralnæringsstoffer som Ca2+, Mg2+, N og P og lavere pH enn kalkfattig jordvannsmyrtorv i samme geografiske område. Dette skyldes at jordvannsmyr tilføres jordvann med oppløste næringsstoffer i tillegg til nedbørvannet. Nedbørsmyr blir likevel ikke blir beskrevet som et trinn 0 langs kalkinnholdsgradienten, det vil si som en enda mer kalkfattig myr enn den mest kalkfattige jordvannsmyra. Det skyldes at det er en kvalitativ, ikke bare en kvantitativ forskjell i torv- og vannkjemi mellom nedbørsmyr og jordvannsmyr. I nedbørsmyrtorv er det dessuten stor variasjon i innholdet av plantenæringsstoffer. Nedbørens kjemiske sammensetning varierer med avstand fra havet og dermed påvirkning fra havvinder som tar med seg sjøsalter. Etter kraftige stormer kan sjøsalteffekten spores mange mil innover land i form av økte Na+ og Cl–-konsentrasjoner i nedbør og avrenning. Nedbørsmyr kan derfor ikke regnes som et spesifikt nederste trinn langs en gradient med pH og Ca2+-konsentrasjon som nøkkelvariabler. Med unntak av ytterste kyststrøk fra Rogaland og nordover er imidlertid nedbørsmyrtorva svært fattig på nitrogen og mineralnæring. Antall karplanter, moser og lav som normalt finnes på nedbørsmyr i Norge, er begrenset til omtrent 40 arter. I nedbørsmyr og fattig jordvannsmyr holder torvmosene tilbake næringsstoffer som karplantene kunne hatt nytte av. Torvmosene fungerer nemlig som ionebyttere ved at H+-ioner på mosenes overflate byttes mot Ca2+ og andre kationer i vannet. Dette begrenser mineralnæringstilgangen til karplantenes røtter. Noen arter har utviklet spesielle tilpasninger til dette ekstremt næringsfattige miljøet, for eksempel soldogg Drosera spp.. Små insekter blir sittende fast i bladenes kjertelhår som også skiller ut enzymer som løser opp fangsten. Denne tilleggsnæringen gjør at de kjøttetende soldoggartene klarer seg på nedbørsmyr. Langs LM-TV Tørrleggingsvarighet domineres mykmattene i Sørøst-Norge av vasstorvmose Sphagnum cuspidatum, fastmattene av rødtorvmose S. rubellum, dvergtorvmose S. tenellum og svelttorvmose S. balticum, og tuene i nederste nivå av rødtorvmose og i øverste nivå av rusttorvmose S. fuscum. I aktivt voksende torv er oftest grensa mellom fastmatter og røsslyngdominerte tuer skarp. Krekling Empetrum nigrum foretrekker det øverste tuenivået (LM-TV_k), mens bjønnskjegg Trichophorum cespitosum er begrenset til øvre fastmatter og nedre tuenivå. Hvitmyrak Rhynchospora alba er konsentrert til mykmatter og nedre fastmatter. Torvull Eriophorum vaginatum og hvitlyng Andromeda polifolia er blant de få vanlige artene som finnes langs hele gradienten fra mykmatte til øvre tuenivå. Lav skyr vanligvis fastmatter og mangler helt i mykmatter, med noen få unntak som for eksempel fnaslav Cladonia squamosa og pigglav C. uncialis. Tuene kan derimot flekkvis domineres av lav som grå reinlav C. rangiferina, kvitkrull C. stellaris og lys reinlav C. arbuscula agg. Nedbørsmyr og kalkfattig jordvannsmyr vil typisk ha lavdominerte områder som alternerer med torvmose- og levermosedominerte flekker i et uregelmessig mønster som kan endre seg i løpet av noen tiår. Verken lav eller levermoser produserer nok organisk materiale til å kunne danne eller akkumulere torv. Over tid vil derfor torvmosedominerte flekker heve seg, mens flekker dominert av de andre artsgruppene vil synke relativt til andre grupper. I løpet av noen tiår kan derfor levermose- og lavdominerte flekker synke fra tuenivå til fastmattenivå. Da dør busklavene og etterlater seg flekker med naken torv. Dynamikken i dette systemet er som en 'stein-saks-papirlek' der artene overtar for hverandre. Torvmosemattene ('papir') infiltreres av levermoser ('saks') som noen ganger tar overhånd, for så å vokse over torvmosehodene og dermed redusere torvmosenes vitalitet. Lav ('stein') invaderer levermoseflekker lettere enn torvmosedominerte flekker. Først når lavmattene etter hvert synker til fastmattenivå, utsettes for hyppig oversvømmelse og etterlater naken torv, ligger det til rette for nykolonisering av torvmoser. Både NA-VA01 Åpen jordvannsmyr og NA-VC01 Åpen nedbørsmyr inneholder variasjon fra myrkantpreget til myrflatepreget natur, men kant på nedbørsmyr mangler fastmatter og mykmatter. I myrkantpreget åpen nedbørsmyr erstattes røsslyng Calluna vulgaris helt eller delvis av bærlyngarter som blokkebær Vaccinium uliginosum, blåbær V. myrtillus og tyttebær V. vitis-idaea. I tillegg er små furuer vanligere, og det blir mer av skogsmoser som furumose Pleurozium schreberi og etasjemose Hylocomium splendens. I nedbørfattige, kontinentale områder med kaldt vinterklima der tuene er høye og lett blåses fri for snø om vinteren, finnes tuevegetasjon som er preget av LM-VI Vindutsatthet, og som inneholder mange arter som kjennetegner hovedtypen NA-TD06 Rabbe (se avsnittet om rabbe-snøleiegradienten). Overgangen mellom minerogen (jordpåvirket) og ombrogen (nedbørpåvirket) torv er mange steder gradvis, både over tid og i rom (se del 3). Noen steder kan overgangen imidlertid være påfallende skarp, for eksempel der klare grenser langs flere miljøvariabler faller sammen. Slike grenser finner vi for eksempel der et hellende, tuedominert høgmyrsegment møter en bløt, kalkrik lagg. Det finner vi først og fremst i store, intakte torvmassiv av TM-H03 Platåhøgmyr og TM-H04 Konsentrisk høgmyr, men velutviklete, intakte torvmassiv av disse typene er sjeldne. De fleste høgmyrer er små og ensidig hellende, med uklar avgrensning mot tilstøtende jordvannsmyrpartier. Når overgangen er uklar, som for eksempel langs kanten av et nesten flatt dråg som gradvis går over i en eksentrisk hvelvet (ensidig hellende) myrflate, varierer artssammensetning gradvis, og arter som krever jordvannstilførsel, minsker også gradvis. Skillet mellom nedbørsmyrtorv og jordvannsmyrtorv er en viktig økologisk grense. Spørsmålet om hvilke arter som kan brukes som indikatorer på jordvannstilførsel, har derfor vært et sentralt tema i fennoskandisk vegetasjonsøkologi i mange tiår. Dette er et vanskelig spørsmål fordi tilført vann fra bakken (fastmarka) ikke kan påvises ved kjemiske, hydrologiske eller andre fysiske målinger. Når overgangen er gradvis, er forskjellen mellom nedbørsmyr og svært kalkfattig jordvannsmyr minimal i alle målbare egenskaper. Selv om torv er et lite gjennomtrengelig substrat, er det dessuten bare i en hvelvet, kuppelformet høgmyr som 'hever seg over terrenget', at det er mulig å utelukke svak jordvannspåvirkning, for eksempel fra grunnvannsfremspring, bekkeløp under torva eller ekstrem flom. Arter med veldig dype røtter kan likevel få tak i næring fra jordvannspåvirket torv i dypere torvlag. Nedbørsmyrtorvas kjemiske sammensetning vil, som nevnt, også variere med avstand fra kysten. I tillegg øker mengden næringsstoffer som tilføres med nedbøren, og dermed blir tilgjengelig for plantene når nedbørmengden øker. Dette gjør det vanskelig å fastslå en grense for tilførsel av vann fra fastmark. Arter kan dessuten ha flaks og lykkes med å etablere seg på et sted utenfor sitt egentlige toleranseområde i en periode med spesielt gunstige forhold, og de kan overleve der i lang tid selv om de ikke er stand til å reprodusere. Bestander av jordvannsmyrarter som flaskestarr Carex rostrata og takrør Phragmites australis kan for eksempel finnes i ombrogen overflatetorv på steder som er under utvikling fra jordvannsmyr til nedbørsmyr. Disse artene kan ha overlevd siden de etablerte seg i jordvannsmyrtorv, blant annet på grunn av et dypt rotsystem som når ned i mer næringsrikt jordvann. En studie av artssammensetningen i sumpskoger i Østmarka øst for Oslo viste at geografisk atskilte sumpskoger med nokså like miljøforhold kan ha svært forskjellig artssammensetning også når de ligger nær hverandre. En mulig forklaring er at hver sumpskog har sin egen unike forstyrrelseshistorikk med vindfall, branner og så videre, og at nye arter etablerer seg i bølger etter slike forstyrrelser fordi marka mesteparten av tiden er dekket av sammenhengende mosematter som hindrer frø i å etablere seg. En slik flekkdynamikk kan være vanlig i våtmarkssystemer med mer eller mindre heldekkende bunnsjikt av vitale moser. Det er fortsatt ikke enighet om hvilke arter som kan brukes som indikatorer på fastmarksvann i ulike deler av Norge. Arter som er sikre indikatorer på jordvannspåvirkning i lavlandet Østafjells, som vortetorvmose Sphagnum papillosum, stivtorvmose S. compactum, duskull Eriophorum angustifolium, klokkelyng Erica tetralix og rome Narthecium ossifragum, blir av mange ansett som indifferente langs vestlandskysten. Flere grundige studier er derfor nødvendige for å komme til bunns i dette. Torvkilder Torvkilder dannes når et kildevannsfremspring over tid dekkes av en torvproduserende mosematte, på samme måte som dannelsen av en gjenvoksningsmyr starter med at flytematter etableres langs bredden av et tjern. Sentralt i kilden kan det finnes et lite 'øye' av åpent vann som holdes åpent av vannstrømmen. En eustatisk kilde er et forutsigbart økosystem. Vanntilgangen er stabil, og vannet er friskt, oksygenrikt og ofte mineralrikt på grunn av løste salter vannet har tatt opp på sin vei gjennom bakken. Kildeøkosystemet er samtidig ekstremt og skiller seg fra naturen omkring ved at substratet er så kaldt. For å trives i en stabil kilde må en art tåle å stå med røttene i kaldt vann hele sommeren. Torvkilder er dermed økosystemer for kuldetolerante arter som ikke liker forandring. Eustatiske torvkilder har en svært karakteristisk artssammensetning med et fåtall dominerende arter. Tette matter av irrgrønne moser er typisk, først og fremst kaldnikke Pohlia wahlenbergii og kildemose-arter Philonotis spp. Dersom kildevannet er kalkrikt, blir kildene mer artsrike, og flere mosearter kommer inn. Flere spesialiserte karplantearter med 'kilde-' i navnet er knyttet til slike steder, for eksempel kildeurt Montia fontana, kildemjølke Epilobium alsinifolium og kildemarikåpe Alchemilla glomerulans. Det kalde miljøet gjør dessuten at arter med utbredelsestyngdepunkt i nordlige fjellstrøk kan finnes i kilder sør for sitt hovedutbredelsesområde. Variasjonen i kildevannsegenskaper er gradvis, og forskjellen mellom topogen (flatt grunnvannsspeil) og soligen (hellende) vanntilførsel er knapt synlig i artssammensetningen. Men med økt friskhet i det soligene vannet øker også innslaget av typiske kildearter. Grensa mellom myr og kilde trekkes mellom basistrinnene KI_c og KI_d, det vil si mellom grunntyper for kildemyr som tilordnes hovedtypen åpen jordvannsmyr, og grunntyper for svak kilde som tilordnes torvkildehovedtypen. I store, stabile torvkilder er det derimot ofte mulig å se variasjon i artssammensetning langs kildevannspåvirkningsgradienten, både fra kildesentrum mot fastmarka omkring kilden og nedstrøms kilden via svak torvkilde og kildemyr til myr uten synlig kildevannspåvirkning. Ikke-torvproduserende våtmarkssystemer NA-VC04 Våtsnøleie er våtmarksparallellen til snøleier på fastmark (NA-TC08). Våtsnøleier forutsetter langvarig snødekke i tillegg til fuktig mark. Vegetasjonen blir derfor også preget av LM-SV Snødekkebetinget vekstsesongreduksjon. Våtsnøleier finnes i fjellet på grusflater som overrisles av smeltevann fra snøfonner som smelter ut sent. Mens det på tilsvarende steder i nedre del av lavalpin bioklimatisk sone ofte finnes myr med grunn torv, blir våtsnøleiene gradvis vanligere mot høyden og dominerer i mellomalpin bioklimatisk sone. Våtsnøleier er også vanlige på Svalbard. Vanntilførselen til våtsnøleier kan ha kildevannspreg. Grensa mot grunnkilde trekkes der vegetasjonen får sterkere preg av kildevannstilførsel enn av langvarig snødekke, det vil si at LM-KI Kildevannspåvirkning dominerer over LM-SV Snødekkebetinget vekstsesongreduksjon. På Svalbard skjer ikke torvdannelse i egentlig forstand, i stedet finnes en myrliknende våtmark på flat mark med permafrost. Denne er såpass spesiell at den tilordnes en egen hovedtype: NA-VC05 Permafrostvåtmark. De øverste 20–30 cm av marka tiner om forsommeren, mens dypere jordlag forblir frosne. Flatt terreng og frossen mark gjør at vannet ikke renner unna. Permafrostvåtmarka er derfor svært våt, store områder har mykmattekarakter, og gjøler er vanlig. Tueliknende partier kan også forekomme. Eventuelt organisk materiale som ikke er brutt ned før det når permafrosten, lagres i permafrostlaget. Permafrostvåtmark kan forekomme sammen med landformen FL-F01 Iskile, som kan danne polygoner. På Svalbard og enkelte steder på fastlandet finnes våtmark på steder preget av sterk frostomrøring av sedimentene og LM-OF Oppfrysing, som resulterer i landformenheten FL-F04 Strukturmark (se del 3). Våtmark med sterkt preg av oppfrysing utgjør hovedtypen NA-VE01 Oppfrysingsvåtmark. Denne typen er betinget av forstyrrelse, i motsetning til de andre spesielle hovedtypene som er nevnt her, som betinges av miljøstress (se del 2). Hevdbetingete våtmarkssystemer Bare jordvannsmyr er egnet som slåttemyr, først og fremst mer kalkrike myrer. Myrslått forutsetter at overflata er jevn og egnet for bruk av ljå. I tillegg må torva være fast nok til å tåle tråkk. Den ideelle slåttemyra er derfor en fastmattedominert, kalkrik myr. Tilrettelegging for slått innebærer fjerning av tuer og eventuelle vedvekster som trær og busker. Langvarig tradisjonell slått kan etter hvert resultere i et parkliknende landskap dominert av homogene fastmatter som likner en velstelt plen. Slått og tilrettelegging for slått har en viss, men ikke vesentlig effekt på artssammensetningen i myr. Starr Carex spp. øker i mengde, det samme gjør noen urter, mens altså vedvekster blir borte. Dersom det finnes torvmoser, går de tilbake, mens myrstjernemose Campylium stellatum ser ut til å gå frem. NA-VK01 Slåttemyr er en hovedtype i NiN til tross for at forskjellen i artssammensetning sammenliknet med NA-VA01 Åpen jordvannsmyr som ikke blir slått, er liten. Dette skyldes prinsippet om at all semi-naturlig mark, det vil si mark med en viss hevdintensitet, skal skilles ut som egne hovedtyper. Mekaniseringen av jordbruket og tilgangen på handelsgjødsel gjorde at slått på myr avtok raskt utover på 1900-tallet. Da annen verdenskrig brøt ut, hadde myrslått opphørt over det meste av landet. Det som finnes av NA-VK01 Slåttemyr i dag, er musealt hevdet slåttemyr der rydding og slått er gjenopptatt for å ta vare på en historisk hevdform i kulturhistorisk restaurerte landskap. Når slåtten opphører, starter en suksesjon mot myr uten preg av hevd. Denne suksesjonen innebærer nyetablering av en tuet overflate og gjengroing med vedvekster. Slåttemyr som ikke forblir åpen, men som etter opphør av slått gror igjen med busker og trær, har sannsynligvis tidligere vært NA-VB01 Myr- og sumpskogsmark som i sin tid ble ryddet. Over tid får artene en mer ujevn fordeling på myra. Selv om slåttemyrpreget på overflatemorfologi og artssammensetning er borte, finnes fortsatt kulturhistoriske spor etter myrslått mange steder i form av utløer og stakkstenger. NA-VK01 Slåttemyr er i prinsippet fortsatt et torvproduserende system selv om fjerning av biomasse reduserer tilfanget av dødt plantemateriale til et minimum så lenge aktiv slått pågår. Mange slåttemyrer har imidlertid også delvis blitt drenert eller på andre måter tilrettelagt for høsting på en måte som har resultert i et semi-naturlig, engpreget våtmarkssystem uten evne til torvproduksjon. Disse samles i hovedtypen NA-VK02 Semi-naturlig våteng, som er den ikke-torvproduserende parallellen til slåttemyr. Myr som brukes til utmarksbeite, får et svakere preg av hevd enn myr som slås. Her er tråkkspor og spredt forekomst av noen beitebegunstigete arter ofte eneste tegn på beitepåvirkning. Først når høyt beitetrykk fra store dyr gir en tråkkpåvirkning som ødelegger torvas struktur, får myra et klart preg av hevd. Store, gjørmedominerte flater uten eller med svært sparsom vegetasjon utgjør hovedtypen NA-VO01 Sterkt tråkkpreget våtmark, som regnes som sterkt endret mark. Annen menneskepåvirkning i våtmarkssystemer Gjennom hele 1900-tallet har myr- og sumpskogsmark blitt grøftet for å fremme produksjon av trevirke. Drenering av myr medfører endring av miljøforholdene i retning fastmark, en langsom prosess som ikke nødvendigvis fører til målet. Grøftet myr tilhører hovedtypen NA-VM01 Sterkt endret torvmark inntil dreneringsprosessen har resultert i et fastmarkssystem." Mer om akrotelmen og katotelmen
Fra Natur i Norge side 147 (se ovenfor): "Torvmosenes årlige lengdevekst bestemmes av voksestedforholdene og kan bli så mye som 40 mm per år i en lavlandsmyr. Dette er imidlertid bare én blant mange faktorer som bestemmer torv-markas høydetilvekst. Allerede noen få centimeter under torvoverflata begynner nedbrytningen av visne torvmoser og annet dødt organisk materiale. Nedbrytningen fortsetter så lenge torvlaget er gjennomluftet, men er minimal under laveste grunnvannsspeilnivå. Derfor går det et viktig skille mellom det øverste, periodevis gjennomluftete torvlaget, akrotelmen, og det permanent vannmettete torvlaget, katotelmen. Mosematerialet blir dessuten stadig presset sammen under den økende tyngden av nytt materiale. Dersom vi ser bort fra nedbrytningen av materiale i katotelmen, bestemmes torvmassivets høydetilvekst av tilfanget av nytt organisk materiale til katotelmen." (Min utheving.) Fra Moen, A., Lyngstad, A. & Øien, D.-I. 2011. Faglig grunnlag til handlingsplan for høgmyr i innlandet (typisk høgmyr). – NTNU Vitensk.mus. Rapp. bot. Ser. 2011-3: 1-60: Hydrologi, akrotelm og katotelmSide 16: "Hydrologi er læren om vannet, og kunnskap om forekomst, fordeling, fysiske og kjemiske egenskaper av vannet i myra er av fundamental betydning for å forstå myras utvikling og økologi. Grunnvann er vannet under grunnvannsspeilet som er det nivået der fritt vann forekommer når det finnes hull i marka. Dybden til grunnvannsspeilet er av største betydning for myrøkosystemene, og ikke minst for høgmyrene. Disse myrene har to typer torvlag (Ivanov 1981): Akrotelmen (acrotelm; mangler norsk betegnelse) er torvlaget som ligger over grunnvannsspeilet. Her er ikke torva vannmettet, og i dette aktive laget med bra tilgang på oksygen foregår viktige prosesser, bl.a. omfattende nedbryting av plantematerialet. Dybden av akrotelmen på et sted i myra varierer med vanntilførselen, for eksempel med nedbøren. I forsenkninger (løsbunn/mykmatte) på høgmyra kan akrotelmen mangle, mens den kan være mer enn 50 cm i de høge tuene og på tresatte myrparti der planterøttene til trær får tilstrekkelig med oksygen. Katotelmen (catotelm; mangler norsk betegnelse) ligger under akrotelmen, dvs. under grunnvannsspeilet. Her er det så godt som anaerobe forhold, og svært sein nedbrytning. I dette inaktive laget er torva vanligvis mer nedbrutt og svartere, og katotelmen utgjør det aller meste av torva på myra. Det er uenighet om differensieringen i akrotelm og katotelm er viktig (gir mening) for alle typer av myrmassiv. Imidlertid gir differensieringen viktig informasjon på typisk høgmyr, og Joosten & Clarke (2002) kaller typisk høgmyr for 'Acrotelm mire'. Etter som dødt plantemateriale akkumulerer på overflata (i akrotelmen) på høgmyr øker også tykkelsen på katotelmen. Selv om nedbrytningen pr. volumenhet er mye større i akrotelmen enn i katotelmen, blir ei grense nådd hvor forskjellen mellom de to lagene er slik at den totale nedbrytingen i katotelmen blir lik med akkumuleringen i akrotelmen. Dette begrenser da tykkelsen som høgmyra kan nå. Om det kommer en klimaforandring (for eksempel økt nedbør) kan veksten fortsette. På denne måten er høgmyra et arkiv for kunnskap om klimaforhold (Ivanov 1981, Ingram 1983). Ei grøft gjennom dette 'byggverket' av torv og vann kan ødelegge hele balansen i systemet, og derved den naturlige utviklingen." Litt om min bakgrunn: Knyttet til NTNU Vitenskapsmuseet fra 1975 til 1984Tor Øystein Olsen jobbet da tett sammen med professor Asbjørn Moen på Vitenskapsmuseet. Les om arbeidet her: NTNU Vitenskapsmuseet. Myr: forskning, forvaltning og overvåking Les om myr på Store norske leksikon. Les om myr på Wikipedia. Les om myr Universitetet i Oslo. Les om Restaurering av Skjervenmyra (Bruamyra) Om sletting av tuedannende arter, tuer og ujevnt bunnsjikt i slåttemyrvegetasjon
|
|
|







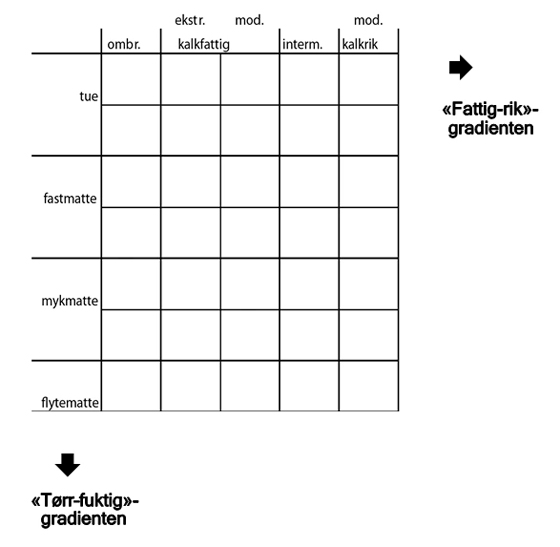










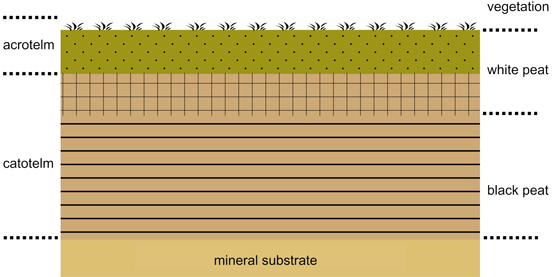
Oversikt over arrangementer:
:Maridalsspillet

:
Årbok Maridalens Venner 2026

Video av Johnny Fagerstrøm 2021:
Film om Maridalen

:
Margaretakirken i fordums drakt video

:
Maridalen. NRK 1972

:
Digitalisert kart over Maridalen
:
Historiske ferdselsveier fra Oslo gjennom Nordmarka til Hakadal og Hadeland

Ansvarlig redaktør: Tor Øystein Olsen. Støtt Maridalens Venners arbeid - kontonr. 0530 58 56349